When salamanders invaded the Dinaric Karst: convergence, history, and reinvention of the troglobitic olm
It is a very good time to be interested in salamanders. Partly because of my reading-up on other areas of direct interest (like cryptic diversity [see previous post], the discovery of new species, declines in global biodiversity, Cretaceous biogeography, radical homoplasy, polymorphism, and evolutionary reversals), I am returning again and again to the salamander literature. Mostly, these studies are about plethodontids, where a lot of really neat research is being done. I will post to the blog about most of this at some stage (if you can’t wait that long see Hanken 1999, Parra-Olea et al. 2001, Chippindale et al. 2004, Mueller et al. 2004 and Min et al. 2005 to see where I’m coming from [update: go here for part I on plethodontids]).
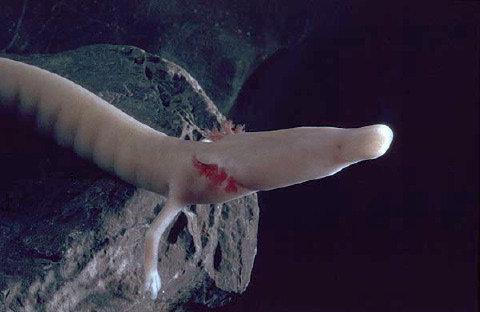
It’s not a plethodontid (it’s a proteid), but one of the most unusual and interesting of amphibians has to be the Olm (Proteus anguinus), an unusual long-bodied cave-dwelling salamander from SE Europe. Olms were the first specialised cave-dwelling animals (so-called stygobionts or troglobites) to be discovered, they were traditionally identified as dragon larvae by local people, and they remain mysterious and the source of controversy, debate and discovery. I’ve had a special affinity for olms since seeing them (live) in the former Yugoslavia in 1987, and after a colleague published a brief article on them in 2004 I ended up compiling and publishing my olm-related thoughts. In the interests of re-cycling that text I reproduce it here, in updated form.
What might be the most fascinating fact concerning olms is the most poorly-known and least mentioned one: the 1986 discovery of a surface-dwelling olm, described in 1994 by Boris Sket and Jan Willem Arntzen. So olms aren’t just ‘unusual long-bodied cave-dwelling salamanders’ – they now exist in two forms, the cave-dwelling White olm Proteus anguinus anguinus and the surface-dwelling Black olm or Brown olm P. a. parkelj. Unlike the unpigmented nominal form with its skin-covered eyes, P. a. parkelj (presently known only from Bela Krajina in SE Slovenia) is dark brown or black and has externally visible (albeit small) eyes. Because White olms produce melanin when kept in sunlight (and are thus not albinistic as sometimes implied), the difference in colour between the two forms is not unexpected. However, there are also other, more important differences separating the two. P. a. parkelj differs from the nominal form in also having a proportionally shorter, broader and more muscular head, fewer teeth, a proportionally longer body and a proportionally shorter tail and limbs (Sket & Arntzen 1994).
Most of the features which distinguish P. a. parkelj from P. a. anguinus are plesiomorphies [= features not unique to olms, but present also in related salamanders] and hence P. a. parkelj may be the ancestor of the White olm. Having said that, one of the most interesting contentions made recently about olms (Sket 1997) is that the different cave-dwelling olm populations may have evolved independently from different ancestral populations. If this is correct it may be that the different White olm populations represent different species which resemble one another by convergent evolution, and which have partly or mostly fused as they have met up within the Dinaric karst system. Sket (1997) thought that morphological and genetic differences observed among olms might provide support for this view and, ironically, if correct it would mean that several old species names proposed for different cave-dwelling olms might need to be resurrected. Fitzinger (1850) named seven new olm species within his genus Hypochthon (H. zoissii, H. schreibersii, H. freyeri, H. haidingeri, H. laurentii, H. xanthostictus and H. carrarae), though given that the type localities for some of them were just a few km apart, it’s unlikely that they really were distinct taxa.
It’s worth saying that olms almost certainly aren’t ancient relicts, or living fossils. In fact, they must be young and recently evolved. Why? During the Pleistocene, the Dinaric area was so close to areas that were fully glaciated that temperature there must have been at or below freezing. This is far too cold for olms, which require temperatures of 6-18ºC for their eggs and larvae to develop (and toward the upper end of that range is best). Furthermore, karstification and the development of underground streams only began in the Dinaric region during the late Pliocene at the earliest, apparently. In view of these problems, olms either (1) survived in surface waters in the region, where summer temperatures were just about tolerable (but where winter temperatures would have made life difficult), or (2) moved into the area from a warmer, southerly refuge (Griffiths 1996, Sket 1997). It isn’t yet known which was the case: more research is needed. Whatever, troglobitic olm populations must have evolved within the last 10,000 years or so, and presumably the specialised troglobitic morphology of living olms evolved during this time. Similarly recent invasions of cave systems appear to have occurred among various troglobitic fishes.
As mentioned earlier, olms were the earliest troglobites to be discovered. While it’s been stated on occasion that the species was discovered as recently as 1875 (Laňka & Vít 1985, Keeling 2004), olms first became widely known in 1689 when Baron Johann Weichard Valvasor wrote about the animals in his book on the Yugoslavian province of Carniola (on which see below). However, it wasn’t until the mid-1700s that the animals become the subject of proper scientific debate. At this time Slovenian scientist Giovanni Scopoli ‘discovered’ olms and realized just how extraordinary they were (Scopoli 1772). He planned to describe the animal scientifically and enhance his reputation by doing so. We know that Scopoli sent pictures of olms to Carl von Linne and that Linne and Scopoli disagreed as to whether the animals were a distinct new genus (Scopoli’s view), or the juveniles of something else (Linne’s view). However, the Austrian anatomist J. N. Laurenti became very interested in olms at the same time as Scopoli (apparently because of a specimen Scopoli had sent to one of the Laurenti’s friends) and, in 1768, published the first scientific description of the species (Laurenti 1768). This is the ‘official’ date of the White olm’s scientific discovery. Laurenti’s choice of generic name for the olm (Proteus) is based on the Greek god Proteus but it is not Proteus’ shape-shifting ability that Laurenti had in mind, but rather his status as shepherd of sea creatures. Laurenti’s work on the olm did not actually become that well known and it was Karl von Schreibers’ work of the 1790s and early 1800s which made olms well known among scientists.
Olm distribution has been the subject of much confusion and speculation. Presently, olms are restricted to the Dinaric Karst, a region that extends from the Soca (formerly the Isonzo) River (near Trieste) in SE Friuli-Venezia Giulia, Italy to the Trebišnica River in eastern Herzogovina. In between Italy and Herzogovina, olms also occur in southern Slovenia, southern Croatia and parts of Bosnia. Little known is that the species has been recorded from localities in France (Moulis) and Germany (Harz). These extralimital records are all apparently due to human introduction however. They are also found in the Parolini Grotto, Vicenza, northern Italy, but their presence here is due to human introduction also. Of further interest, the locality mentioned by Valvasor (1689) – the spring of Lintvern, near Vrhnika – is actually outside of the Dinaric Karst, and is unlike the other areas inhabited by olms in geology and geomorphology. It seems that Valvasor made the logical (but incorrect) assumption that Lintvern (which is a garbled form of the German word Lindwurm, meaning dragon) was so named because it was the source of olms (which were fancifully regarded as dragon larvae at the time).
Keeling (2004) implied that Carniola (note: not Carinola) might be the Italian part of the olm’s range and also wondered if Carniola might still be part of Austria. Carniola is today called Kranjska and was controlled by Austrian royalty until 1918 (consequently, the ruling classes there spoke German until the 20th century). It is today part of central Slovenia and is thus not either the Italian part of the olm’s range, nor an Austrian extension of the species’ range.
Bizarrely, olms were traded during Victorian times as exotic pets and were apparently available in Britain as such (which raises the remote possibility that they might have been introduced to British cave systems in the same way that they were in French, German and Italian ones). During the 1950s it was reported that olms were present in the Carpathian karst of eastern Serbia, and in 1960 a team of speleologists from Ljubljana led an expedition to the region to investigate this possibility. They didn't find any olms there, and nor has anyone else since.
Olms have been horrendously over-collected for scientific use and were also apparently collected by farmers for use as pig food. One of the greatest problems facing olms today is metal poisoning caused by industrial pollution and a number of populations have declined as a result of such. P. a. parkelj is under strict legal protection. Olms have been protected in Slovenia at least since 1949 and elsewhere in their range they are widely recognized as deserving protection.
Finally, regarding diet and breeding, olms apparently mostly detect their prey using chemical clues and the detection of water currents but they also possess electroreceptive organs in the head and thus presumably employ electroreception. Despite their vestigial nature, the eyes of White olms are not completely useless and are able to detect light. Olms appear to mostly prey on aquatic crustaceans but also eat snails and insect larvae. Captive specimens have eaten worms and adults may be cannibalistic on occasion. When I visited Postojina we were told that the olms on display were not fed both because their food proved hard to procure, and because they were quite able to survive for years without feeding. Indeed there was apparently a specimen kept at the Faculty of Biotechnology in Ljubljana which survived for an astonishing 12 years without food. Olms are long-lived, reaching sexual maturity between 7 and 14 years, and almost certainly ordinarily live for more than 50 years, though ages twice this have been suggested by some writers.
PS - for the latest news on Tetrapod Zoology do go here.
The photo above is from here.
Refs - -
Chippindale, P. T., Bonett, R. M., Baldwin, A. S. & Wiens, J. J. 2004. Phylogenetic evidence for a major reversal of life-history evolution in plethodontid salamanders. Evolution 58, 2809-2822.
Fitzinger, L. 1850. Ueber den Proteus anguinus der Autoren. Sitz.-Ber. Akad. Wiss., Math.-naturw. Cl. 5, 291-303.
Griffiths, R. A. 1996. Newts and Salamanders of Europe. T & A D Poyser (London).
Hanken, J. 1999. Why are there so many new amphibian species when amphibians are declining? Trends in Ecology & Evolution 14, 7-8.
Keeling, C. 2004. Olm. Mainly About Animals July 2004, 20-21.
Laňka, V. & Vít, Z. 1985. Amphibians and Reptiles. Hamlyn (London).
Laurenti, J. L. 1768. Specimen Medicum Exhibens Synopsis Reptilium Emendatum. Joan. Thomae (Vienna).
Min, M. S., Yang, S. Y., Bonett, R. M., Vieites, D. R., Brandon, R. A. & Wake, D. B. 2005. Discovery of the first Asian plethodontid salamander. Nature 435, 87-90.
Mueller, R. L., Macey, J. R., Jaekel, M., Wake, D. B. & Boore, J. L. 2004. Morphological homoplasy, life history evolution, and historical biogeography of plethodontid salamanders inferred from complete mitochondrial genomes. Proceedings of the National Academy of Sciences 101, 13820-13825.
Parra-Olea, G. & Wake, D. B. 2001. Extreme morphological and ecological homoplasy in tropical salamanders. Proceedings of the National Academy of Sciences 98, 7888-7891.
Scopoli, J. A. 1772. Annus Quintus Historico-Naturalis. C. G. Hilscher (Lipsiae).
Sket, B. 1997. Distribution of Proteus (Amphibia: Urodela: Proteidae) and its possible explanation. Journal of Biogeography 24, 263-280.
- . & Arntzen, J. W. 1994. A black, non-troglomorphic amphibian from the karst of Slovenia: Proteus anguinus parkelj n. ssp. (Urodela: Proteidae). Bijdr. Dierk. 64, 33-53.
Valvasor, J. W. 1689. Die Ehre des Herzogthums Crain. W. M. Endtner (Nuernberg).
posted by Darren Naish at 11:22 PM
![]()
![]()


4 Comments:
Curiously I had just finished reading about the plethodontid salamander Eurycea tynerensis:
Ronald M Bonett & Paul T Chippindale. Streambed microstructure predicts evolution of development and life history mode in the plethodontid salamander Eurycea tynerensis. BMC Biology, 4:6 doi:10.1186/1741-7007-4-6
http://www.biomedcentral.com/1741-7007/4/6/abstract
But these guys are neat also. Thanks for sharingthis.
Thanks for the heads up - I'll check this paper out, thanks. Sounds interesting. It certainly seems that plethodontids are remarkably 'plastic' in terms of the evolutionary things they can do. But are they really more so than other groups, or it is just because lots of really neat, cutting-edge stuff is being done on them?
Earlier today I saw the name Dinaric and thought, where have I seen that before? The answer (of course) was this post, and the place I saw Dinaric mentioned earlier was in Dalla Vecchia's chapter in The Thunder-Lizards about sauropods from Early Cretaceous carbonate platforms...which erode into karsts, and pick up bizarre salamander taxa. What a weird world.
Yes - the Cretaceous dinosaurs and other taxa that Dalla Vecchia and others have worked on are from the so-called ADCP (Adriatic-Dinaric Carbonate Platform). In the Cretaceous it was variously submerged, an island, or an archipelago. Looks like the dinosaurs that lived there were unusual island endemics.
Post a Comment
<< Home