Basal tyrant dinosaurs and my pet Mirischia
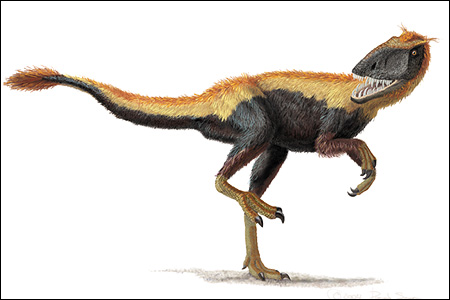 I knew when I started this blog that my posts would be a random compilation of thoughts and observations on both living and fossil tetrapods, and I’d somehow imagined that the posts about fossil tetrapods would draw more attention than those on the living ones. I felt this prediction had come true when the post on azhdarchid pterosaurs attracted a record 14 responses.
I knew when I started this blog that my posts would be a random compilation of thoughts and observations on both living and fossil tetrapods, and I’d somehow imagined that the posts about fossil tetrapods would draw more attention than those on the living ones. I felt this prediction had come true when the post on azhdarchid pterosaurs attracted a record 14 responses.But by and large the posts on extant animals have drawn equal amounts of praise and attention. I’m a vertebrate palaeontologist specializing in dinosaurs, but I’m as interested in living animals as I am in long-dead ones, so this is all fine by me. I do sometimes get paranoid that I’m not bigging up my own subject enough, however, and for this reason I feel pressured to produce a post on dinosaurs. And given that I have my proverbial fingers on the pulse of basal tyrant dinosaur research, they’re as good a group to examine as any.
A few introductory comments for novices before I begin. Theropoda is the group name for the predatory dinosaurs (including birds), and Coelurosauria is a major theropod group that includes birds and all the bird-like theropods (including tyrannosauroids). Tyrannosauroidea includes the familiar giant tyrannosaurs like Tyrannosaurus of the Upper Cretaceous as well as an assortment of less familiar theropods, the oldest of which are from the Upper Jurassic.
So is it time to produce the definitive blog post on Eotyrannus lengi, the dinosaur I did my phd on? Maybe. Actually, no. Eotyrannus was named by myself and colleagues in 2001, and in that initial paper we proposed that it was a basal tyrannosauroid, and one of the most basal members of the group (Hutt et al. 2001). Since then I’ve described the anatomy of Eotyrannus in full and tedious detail (the relevant thesis chapter is 118 pp and over 30,000 words long) and have come to know it well. I am now utterly convinced that it is a tyrannosauroid, and the results of my cladistic analysis (and those of others – see Holtz 2004) support this. Every dinosaur expert who knows anything about Eotyrannus agrees, by the way.
It turns out that the 2001 characterisation of Eotyrannus is horrendously wrong, as a new rigorous skeletal reconstruction (to be published soon) shows. The animal looked substantially different from the way I initially reconstructed it (see Naish 2001, Naish et al. 2001 and Holtz 2004), and in detail it’s proved to be strikingly odd and unique in many, many features. I’ll talk about these details some time soon, but not now. In fact I’m currently making arrangements to get the full monograph published in a high prestige journal, and with the co-authorship of a leading expert on tyrant dinosaurs I hope to produce an important work on tyrannosauroid phylogeny and morphology. More on this as and when it happens.
Since Eotyrannus was published a few very interesting things have been happening in the world of basal tyrannosauroids. I was aiming to discuss all of these here, but as usual I veered off at a tangent and have hardly scratched the surface. Eotyrannus seems to have been a mid-sized theropod. The type specimen is a juvenile individual that would have been 4.5 m long when complete, but fragmentary specimens from larger individuals indicate that adults were perhaps around 7 m long. But other basal tyrannosauroids are way smaller than this, mostly being less than 3 m long.
We begin with Dilong paradoxus, a basal tyrannosauroid known from excellent near-complete specimens from the Lower Cretaceous Yixian Formation of Liaoning Province, China (Xu et al. 2004). The Yixian Formation is the now famous unit that has produced all those little coelurosaurs with feathers and other integumentary structures preserved, and Dilong is no exception. It is preserved with simple quill-like integumentary structures that seem antecedent to the true complex feathers that evolved later.
Dilong has proved to be a sort of Rosetta stone for me, allowing several previously enigmatic Lower Cretaceous coelurosaurs to be reinterpreted as additional basal tyrannosauroids. Given that I haven’t published the relevant details, I’d be silly if I gave the game away here, but to be honest the only people who really care about this already know the relevant details, and furthermore I am silly anyway. The news is that a controversial little coelurosaur from the Isle of Wight’s Wessex Formation, Calamosaurus foxi (known only from two cervical vertebrae, one of them incomplete), is so similar to the cervical vertebrae of Dilong that I am confident that it too should be identified as a basal tyrannosauroid. This is mentioned in a large manuscript that came back from review some weeks ago and is currently undergoing revision, but the full story is to be revealed in a short paper that’s been completed and reviewed but now awaits post-review revamping.
I previously had Calamosaurus down as a compsognathid (Naish et al. 2001). Compsognathids are, like tyrannosauroids, basal coelurosaurs, but they retained small body size throughout their history (so far as we know). They were also conservative in all being rather alike: morphologically unspecialized with relatively short forelimbs, rather long and gracile legs and feet, and a long tail. There’s a lot more that could be said about them but this isn’t the time. Here’s the thing: the fact that Calamosaurus probably isn’t one of them after all leads us to a key question. Namely, do all the other compsognathids really go together, or is Compsognathidae as currently perceived actually an artificial assemblage of distantly related (yet superficially similar) theropods?
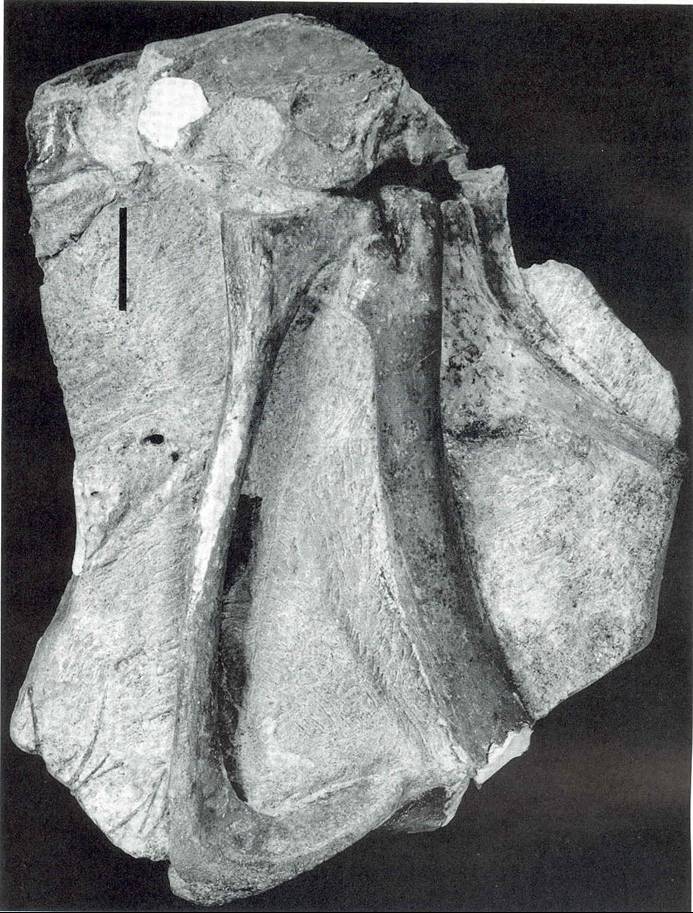 Back in 2004 I and colleagues named the new Brazilian theropod Mirischia asymmetrica (photo at left). That name means ‘asymmetrical wonderful pelvis’, and I think it’s a rather good descriptive name. The only known specimen really is a ‘wonderful pelvis’ (though it also includes some of the hindlimb bones), as it is 3-D and fantastically well-preserved, including even soft tissues like part of the gut and a probable post-pubic air sac (and to know the importance of that latter feature you’ll have to wait for a future post). As you might guess, it’s also asymmetrical, but I won’t talk about that here. I also don’t want to talk about the embarrassing fact that Mirischia’s generic name might imply that it was named after the fund-giving Mirisch Foundation, but (annoyingly) I only found this out after the publication of the Mirischia paper.
Back in 2004 I and colleagues named the new Brazilian theropod Mirischia asymmetrica (photo at left). That name means ‘asymmetrical wonderful pelvis’, and I think it’s a rather good descriptive name. The only known specimen really is a ‘wonderful pelvis’ (though it also includes some of the hindlimb bones), as it is 3-D and fantastically well-preserved, including even soft tissues like part of the gut and a probable post-pubic air sac (and to know the importance of that latter feature you’ll have to wait for a future post). As you might guess, it’s also asymmetrical, but I won’t talk about that here. I also don’t want to talk about the embarrassing fact that Mirischia’s generic name might imply that it was named after the fund-giving Mirisch Foundation, but (annoyingly) I only found this out after the publication of the Mirischia paper.Anyway, Mirischia is enough like Compsognathus – particularly in the detailed anatomy of its pubis – for me to convince myself that it must belong together with Compsognathus in a little clade, and by definition this clade has to be called Compsognathidae. So in the 2004 paper, Naish et al. argued strenuously that (1) there is a little clade called Compsognathidae, the members of which can be united on the basis of shared derived characters, and (2) in addition to Compsognathus, this clade includes Sinosauropteryx and Mirischia. This published opinion is in part the result of a long series of to-ings and fro-ings between myself and my good friend Nick Longrich, for Nick has long been a vocal opponent of the idea that Compsognathidae really is monophyletic. Nick thinks that some so-called compsognathids are basal maniraptorans, and that others aren’t even coelurosaurs, but to date he’s only published an abstract on this (and to see why this is interesting and important you’ll have to wait until yet another future post. Think alvarezsaurids and stagodontids). Alas poor Longrich, surely he can’t be right. By the way, if you’re at all interested in what Nick looks like, click here. Sorry Nick.
 Here’s where we bring Dilong back in. The pelvic anatomy of Dilong – an undoubted basal tyrannosauroid – is (like that of Mirischia) rather like that of Compsognathus. This is disturbing, as it all but destroys the reasons for thinking that Mirischia can only be a compsognathid. Might it actually be a basal tyrannosauroid? This has implications for another supposed compsognathid: the Isle of Wight taxon Aristosuchus pusillus (photo at left, with interpretative restoration below) which, again, I’d previously identified with confidence as a close relative of Compsognathus (Naish 2002, Naish et al. 2001, 2004). So we can now doubt that those ‘compsognathids’ known only from pelvic material really truly are compsognathids. Again, this is an idea that I’m mentioning in that large in-preparation manuscript.
Here’s where we bring Dilong back in. The pelvic anatomy of Dilong – an undoubted basal tyrannosauroid – is (like that of Mirischia) rather like that of Compsognathus. This is disturbing, as it all but destroys the reasons for thinking that Mirischia can only be a compsognathid. Might it actually be a basal tyrannosauroid? This has implications for another supposed compsognathid: the Isle of Wight taxon Aristosuchus pusillus (photo at left, with interpretative restoration below) which, again, I’d previously identified with confidence as a close relative of Compsognathus (Naish 2002, Naish et al. 2001, 2004). So we can now doubt that those ‘compsognathids’ known only from pelvic material really truly are compsognathids. Again, this is an idea that I’m mentioning in that large in-preparation manuscript.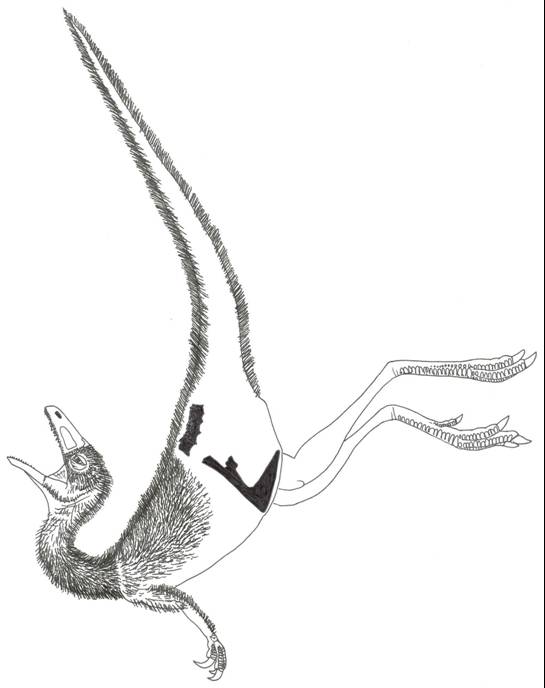 In my thesis I tried to test all of this: I included all of the relevant taxa (except Aristosuchus, as I decided it wasn’t complete enough to code) and all of the characters that have been used in this debate. And the result? Well, yes, there was a monophyletic Compsognathidae, but it consisted only of Compsognathus and one other taxon (and I’d rather not say which taxon that was right now). It wasn’t Mirischia, as this came out as… a basal tyrannosauroid. Two characters helped pull Mirischia into Tyrannosauroidea. One of these was discussed in the Naish et al. (2004) paper but the other was previously overlooked. I’ll not mention them here for fear of giving away all the secrets.
In my thesis I tried to test all of this: I included all of the relevant taxa (except Aristosuchus, as I decided it wasn’t complete enough to code) and all of the characters that have been used in this debate. And the result? Well, yes, there was a monophyletic Compsognathidae, but it consisted only of Compsognathus and one other taxon (and I’d rather not say which taxon that was right now). It wasn’t Mirischia, as this came out as… a basal tyrannosauroid. Two characters helped pull Mirischia into Tyrannosauroidea. One of these was discussed in the Naish et al. (2004) paper but the other was previously overlooked. I’ll not mention them here for fear of giving away all the secrets.Furthermore, other supposed compsognathids did not group with Compsognathidae proper. Instead they were scattered about the base of Coelurosauria. So right now – while further work and further testing and further incorporation of data is needed – I am thinking that Nick was right, and that Compsognathidae in its old, inclusive sense is an artificial grouping.
People sometimes ask what sort of relevance stuff like this really has for our understanding of animals and their evolution. Well, it actually tells us an awful lot of stuff about patterns and trends. If so-called compsognathids – all of them relatively small, ecologically and morphologically generalized, long-limbed, long-tailed theropods that hunt small vertebrate prey – are not a clade but are actually scattered about the base of the coelurosaur family tree, this likely indicates that this ecotype was the ancestral one for coelurosaurs. We might already have thought that based on other lines of evidence, but this would help confirm it. There are indications that ‘compsognathids’ could make a living just about anywhere (for reasons that, again, I’ll have to cover in another post), and if this is valid then again we have another really interesting discovery about evolution at the base of Coelurosauria.
And I’ll have to stop there. I was planning to discuss the Jurassic basal tyrannosauroids Aviatyrannis (from Portugal) and Guanlong (from China). Another time. To readers who already knew all of this stuff, I apologise. To those who didn’t: welcome to the fantastic world of dinosaurs! More to come. For the latest news on Tetrapod Zoology do go here.
Refs - -
Holtz, T. R. 2004. Tyrannosauroidea. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds) The Dinosauria, Second Edition. University of California Press (Berkeley), pp. 111-136.
Hutt, S., Naish, D., Martill, D. M., Barker, M. J. & Newbery, P. 2001. A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England. Cretaceous Research 22, 227-242.
Naish, D. 2001. Eotyrannus lengi, a new coelurosaur from the Isle of Wight. Dino Press 5, 82-91.
- . 2002. The historical taxonomy of the Lower Cretaceous theropods (Dinosauria) Calamospondylus and Aristosuchus from the Isle of Wight. Proceedings of the Geologists’ Association 113, 153-163.
- ., Hutt, S. & Martill, D. M. 2001. Saurischian dinosaurs 2: Theropods. In Martill, D. M. & Naish, D. (eds) Dinosaurs of the Isle of Wight. The Palaeontological Association (London), pp. 242-309.
- ., Martill, D. M. & Frey, E. 2004. Ecology, systematics and biogeographical relationships of dinosaurs, including a new theropod, from the Santana Formation (?Albian, Early Cretaceous) of Brazil. Historical Biology 16, 57-70.
Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q. & Jia, C. 2004. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431, 680-684.
Labels: archosaurs, British dinosaurs, dinosaurs, theropods
posted by Darren Naish at 1:13 AM
![]()
![]()


4 Comments:
So much confusion! The reason it's so hard to sort these creatures into clades is the problem of basal, unspecialized forms, isn't it? They all look sort of like their common ancestor.
It also suggests there was something about Cretaceous ecosystems that welcomed a small, generalized theropod.
I know of various examples of specializations evolving again and again (like the way saber-tooths have turned up in multiple carnivore lineages). There seems to be less attention to recurring generalists. Hm.
Doug M.
I may not, perchance, ask about Juravenator?
Like I said David, I don't want to give out ALL of the secrets. Juravenator is particularly interesting in this mess: details later.
Post a Comment
<< Home