Why putting your hand in a peccary’s mouth is a really bad idea
 In a recent post I discussed the apparent recent discovery of a new, fourth peccary species (Meet peccary # 4). While this case has now become relatively well known in the cryptozoological world, I’ve learnt from Matt Bille* that not all artiodactyl experts are ready to accept the animal, the so-called Giant peccary, as a valid new species. We await developments. But of course there’s lots more to peccaries than their species-level diversity, and here we’re going to look at a few other areas of interest: their former diversity and fossil history, and their fascinating cranial anatomy.
In a recent post I discussed the apparent recent discovery of a new, fourth peccary species (Meet peccary # 4). While this case has now become relatively well known in the cryptozoological world, I’ve learnt from Matt Bille* that not all artiodactyl experts are ready to accept the animal, the so-called Giant peccary, as a valid new species. We await developments. But of course there’s lots more to peccaries than their species-level diversity, and here we’re going to look at a few other areas of interest: their former diversity and fossil history, and their fascinating cranial anatomy.* Best known in the cryptozoology world for the newsletter he previously produced (Exotic Zoology) and for his books: Rumors of Existence and the new Shadows of Existence (see Hancock House website).
Peccaries today are entirely American, so you might be surprised to hear that fossil evidence indicates that they originated in the Old World, and in fact were still there until as recently as the Late Miocene. The oldest named peccary is Egatochoerus* from the Upper Eocene of Thailand (Ducrocq 1994), and similarly-aged taxa are known from southern China. If these forms are peccaries (read on), then they migrated into North America early in their history, as peccaries made their American debut during the Early Oligocene, and possibly even in the Late Eocene given that there are possible records of Thinohyus from this time.
* Incidentally this is one of those taxa whose name incorporates an acronym: in this case EGAT, the Electricity Generating Authority of Thailand.
 At about the same time, peccaries also seem to have got from Asia into Europe as several Oligocene genera – including Doliochoerus and Propalaeochoerus – appeared suddenly in western Europe, yet without antecedents. European peccaries, mostly belonging to the clade Doliochoerinae Simpson, 1945, persisted until the end of the Miocene, and members of this group also inhabited Asia during the Miocene. It was also in the Miocene that peccaries invaded Africa. Morotochoerus from Middle Miocene Kenya seems to be ancestral to Schizochoerus (Pickford 1988), a genus that first appeared in Africa but had spread to Asia and Europe by the Late Miocene (Pickford 1978). Africa-Arabia was an island until the Early Miocene, incidentally, when a contact with Asia was formed, allowing African taxa to colonise Eurasia. This is known as the Proboscidea Datum Event as, obviously, it’s the point when African proboscidean groups make their first appearance in Eurasia. The temporal and geographical distribution of Morotochoerus and Schizochoerus indicates that peccaries invaded Africa from Asia during or after the Proboscidea Datum Event, with one lineage then leaving Africa later in the Miocene. In and out of Africa, in other words (which, incidentally, is the title of a paper on primate evolution by C.-B. Stewart and T. R. Disotell).
At about the same time, peccaries also seem to have got from Asia into Europe as several Oligocene genera – including Doliochoerus and Propalaeochoerus – appeared suddenly in western Europe, yet without antecedents. European peccaries, mostly belonging to the clade Doliochoerinae Simpson, 1945, persisted until the end of the Miocene, and members of this group also inhabited Asia during the Miocene. It was also in the Miocene that peccaries invaded Africa. Morotochoerus from Middle Miocene Kenya seems to be ancestral to Schizochoerus (Pickford 1988), a genus that first appeared in Africa but had spread to Asia and Europe by the Late Miocene (Pickford 1978). Africa-Arabia was an island until the Early Miocene, incidentally, when a contact with Asia was formed, allowing African taxa to colonise Eurasia. This is known as the Proboscidea Datum Event as, obviously, it’s the point when African proboscidean groups make their first appearance in Eurasia. The temporal and geographical distribution of Morotochoerus and Schizochoerus indicates that peccaries invaded Africa from Asia during or after the Proboscidea Datum Event, with one lineage then leaving Africa later in the Miocene. In and out of Africa, in other words (which, incidentally, is the title of a paper on primate evolution by C.-B. Stewart and T. R. Disotell).Some of these Old World peccaries were rather unusual. The doliochoerine Lorancahyus of Miocene Spain, for example, had tubulidentate teeth, and if that word sounds familiar that’s probably because you’ve seen it before as the group name for aardvarks: Tubulidentata. Tubulidentate teeth are traversed by tubules, hence the name, and they’re thought to be an evolutionary response to the ingestion of large abrasive particles (such as quartz grains) such as those taken into the mouth with soil. It’s even been suggested that some fossil teeth from Miocene Spain, originally described as belonging to aardvarks, may actually be from this lineage of peccaries (Pickford & Morales 1998). What were these peccaries doing with their aardvark-like teeth? As usual, we don’t know.
But are these Old World peccaries really peccaries? Well, perhaps not, as some workers have expressed scepticism, stating for example that the identification of Old World peccaries as peccaries proper is ‘not based on synapomorphy’ (Wright 1998, p. 389), the implication therefore being that these taxa might be suids that are convergently peccary-like in a few features. Van der Made (1997) proposed that Old World peccaries are in fact different enough from peccaries proper to be regarded as a different family, Palaeochoeridae Matthew, 1924. This is problematical however as the type genus for this group, Palaeochoerus from Oligocene-Miocene Africa and Europe, almost certainly is a suid proper and not a peccary, Old World or otherwise. Furthermore, Pickford (1998) argued that Van der Made’s concept of Palaeochoeridae is probably polyphyletic, including not only Old World peccaries and suids but also groups like the sanitheriids of the Old World Miocene. On the other hand other workers have described some of the Old World peccaries as ‘unambiguous Tayassuidae’ (Ducrocq 1994, p. 765).
While the debate isn’t over, Old World peccaries do at least share characters with unambiguous American peccaries that aren’t seen in suids: a vertical lower canine located close to the premolar row, a prominent trigonid on the fourth upper premolar, thin enamel, and other features (Pickford & Morales 1989). Old World and New World peccaries also lack many characters present in all suids, but these carry less weight as, theoretically, they could be retained plesiomorphies if Old World peccaries are suids and not peccaries proper. They include the lack of dentary symphyseal splaying around the canines and the lack of an obliquely oriented lower tooth row. But for the time being I’d say that Old World peccaries really do look like peccaries after all, not that I’m an expert.
Like pigs, peccaries use a specialised rhinarial disk for rooting in soil and their snout is specialised for this behaviour. The disk itself is an unusual novelty, the snout is proportionally lengthened compared to that of other artiodactyls and the nuchal muscles (which help support the head) are hypertrophied and with enlarged attachment areas. However, the muscles that operate the disk originate from different points in peccaries and suids.
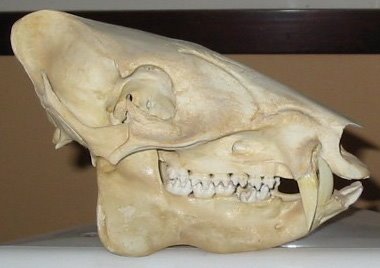 Strangely, all of the skull sutures in adult peccaries are completely closed, and in fact even juveniles exhibit closure of certain of the sutures ordinarily open in young mammals. This obviously rules out the possibility of any sort of cranial kinesis. A research team led by Katherine Rafferty and Susan Herring have been looking at strain patterns in pig skulls, and they’ve found that during occlusion of the teeth, the snout bones are deformed and pull apart slightly at their sutures (Rafferty et al. 2003). I’m guessing that this is somehow relevant to the major fusion of the sutures seen in peccaries (maybe peccaries have evolved a novel solution to coping with strains built up during tooth occlusion), but I don’t know if this area has been studied. It’s surely relevant to stuff we’ll get to in a minute.
Strangely, all of the skull sutures in adult peccaries are completely closed, and in fact even juveniles exhibit closure of certain of the sutures ordinarily open in young mammals. This obviously rules out the possibility of any sort of cranial kinesis. A research team led by Katherine Rafferty and Susan Herring have been looking at strain patterns in pig skulls, and they’ve found that during occlusion of the teeth, the snout bones are deformed and pull apart slightly at their sutures (Rafferty et al. 2003). I’m guessing that this is somehow relevant to the major fusion of the sutures seen in peccaries (maybe peccaries have evolved a novel solution to coping with strains built up during tooth occlusion), but I don’t know if this area has been studied. It’s surely relevant to stuff we’ll get to in a minute.Peccaries are well known for having big, scary canines and, unlike suids, both the upper and lower canines of peccaries are used in biting. Also in contrast to those of suids, both canines in peccaries are vertically implanted: in suids the upper canine exits the maxilla anterolaterally, and then curves dorsally. In all placental mammals the lower canine bites ‘ahead’ of the upper canine (look at sloths and you’ll see that they differ – almost certainly because one of their canines isn’t actually a true canine), and in peccaries the almost total lack of enamel on the posterior surface of the lower canine means that it is constantly sharpened as it moves against the enamelled anterior face of the upper canine. Because the upper and lower canines tightly interlock, peccaries are virtually incapable of moving the lower jaw from side-to-side when the jaws are closed. A special mucosal pocket, bordered by a raised boss on the maxilla termed the canine buttress, houses the lower canine when the jaws are closed (Herring 1972).
The tight interlocking of the canines prevents jaw movement during full occlusion, but there’s more: bony stops around the jaw joint further prevent anteroposterior movement of the jaws when they’re closed. Consequently the jaws can only open and close in a simple hinge-like arrangement. Herring (1972, p. 502) suggested that ‘this action probably helps to guide the lower canine into its correct occlusal relationship, thus preventing injury to soft tissues’.
 But exactly why do the canines interlock? Inspired by Herring’s study, Kiltie (1981) studied peccary teeth in detail and noted a correlation between tooth morphology and a diet of hard nuts and seeds. Kiltie didn’t propose that the canines were used to break open the food items, but that, like the bony stops around the jaw joint itself, their interlocking helped prevent dislocation of the joint when tremendous force was applied across the molar teeth. Several other features of the peccary dentition are in agreement with the idea that peccaries are specialised for breaking open rock-hard objects, as are behavioural studies. Captive animals are reported to often try to break open excessively hard nuts and seeds. I wonder if anyone has ever measured the bite strength of a peccary. Whatever, all the more reason not to put your hand in a peccary’s mouth, and to have those new fences up at Marwell Zoo (go here).
But exactly why do the canines interlock? Inspired by Herring’s study, Kiltie (1981) studied peccary teeth in detail and noted a correlation between tooth morphology and a diet of hard nuts and seeds. Kiltie didn’t propose that the canines were used to break open the food items, but that, like the bony stops around the jaw joint itself, their interlocking helped prevent dislocation of the joint when tremendous force was applied across the molar teeth. Several other features of the peccary dentition are in agreement with the idea that peccaries are specialised for breaking open rock-hard objects, as are behavioural studies. Captive animals are reported to often try to break open excessively hard nuts and seeds. I wonder if anyone has ever measured the bite strength of a peccary. Whatever, all the more reason not to put your hand in a peccary’s mouth, and to have those new fences up at Marwell Zoo (go here).If Kiltie is right about the bracing function of the canines, then this would explain why – unlike many other mammals that use their canines as offensive weapons – peccaries are not sexually dimorphic in canine size. They aren’t dimorphic in body size, nor in head shape, either. But this isn’t the whole story, as fossil peccaries ordinarily are dimorphic, with many forms exhibiting significant (i.e., distinctly bimodal) differences in canine size (Wright 1998). In fact what’s almost bizarre is that fossil populations of the living species exhibit sexual dimorphism in canine size, meaning that the living populations lost dimorphism somewhere along the way.
So what gives? We don’t know why sexual dimorphism was lost in the group (decreasing need to avoid niche overlap, due to declining diversity in contemporaneous megafauna?), but comparison with related groups, and examination of the peccary fossil record, indicates that sexual dimorphism in canine size is primitive for the group (it’s the condition they inherited from their ancestors). Use of the canines as bracing structures therefore looks like an exaptation: a new use for a set of structures that were previously used for something else.
I did want to talk about the history of peccaries in the Americas, but I’ve run out of time. Coming next: probably those sexy tupuxuarids (go here for teaser). For the latest news on Tetrapod Zoology do go here.
Many thanks to Steve Bodio (of Querencia) for the Collared peccary Tayassu tajacu skull photos that appear here. The Collared peccary photo at top is from birdfotos.com and the White-lipped peccary T. pecari photo is from the Cites sites.
Refs - -
Ducrocq, S. 1994. An Eocene peccary from Thailand and the biogeographical origins of the artiodactyl family Tayassuidae. Palaeontology 37, 765-779.
Herring, S. W. 1972. The role of canine morphology in the evolutionary divergence of pigs and peccaries. Journal of Mammalogy 53, 500-512.
Kiltie, R. A. 1981. The function of interlocking canines in rain forest peccaries (Tayassuidae). Journal of Mammalogy 62, 459-469.
Pickford, M. 1978. The taxonomic status and distribution of Schizochoerus (Mammalia, Tayassuidae). Tertiary Research 2, 29-38.
- . 1998. A new genus of Tayassuidae (Mammalia) from the Middle Miocene of Uganda and Kenya. Annales de Paléontologie 84, 275-285.
- . & Morales, J. 1989. On the tayassuid affinities of Xenohyus Ginsburg, 1980, and the description of new fossils from Spain. Estudios Geologicos 45, 233-237.
- . & Morales, J. 1998. A tubulidentate suiform lineage (Tayassuidae, Mammalia) from the Early Miocene of Spain. Comptes Rendu de l’Academie des Sciences, Paris, Serie II 327, 285-290.
Rafferty, K. L., Herring, S. W. & Marshall, C. D. 2003. Biomechanics of the rostrum and the role of facial sutures. Journal of Morphology 257, 33-44.
Van der Made, J. 1997. Systematics and stratigraphy of the genera Taucanamo and Schizochoerus and a classification of the Palaeochoeridae. Proceedings of the Koninkliijke Nedderlandse Akademie voor Wetenschappen 100, 127-139.
Wright, D. B. 1998. Tayassuidae. In Janis, C. M., Scott, K. M. & Jacobs, L. L. (eds) Evolution of Tertiary Mammals of North America. Volume 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Cambridge University Press, pp. 389-401.
posted by Darren Naish at 9:56 PM
![]()
![]()


8 Comments:
You wrote, "In all placental mammals the lower canine bites ‘ahead’ of the lower canine". Did you mean, "The UPPER canine bites 'ahead' of the lower canine"?
Silly mistake, now corrected. Thanks for pointing it out. Lower canines bite 'ahead' of upper canines.
Thanks for the explanation of the peccary's unusual dentition.
Why can't it be that peccary have their dentition because they can, assuming limited lateral motion was already present? Are canines so important for that? I'm imagining an experiment where one removes some peccary canines and sees if it still can crack nuts.
It would be nice to know the kind of stresses we are talking about, but from your comments I infer no one has done that.
Filipe: thanks for your comment. As mentioned in the post, phylogenetic bracketing indicates that peccaries retained big canines from their ancestors, so the proposed assistant bracing function of the canines must have evolved as an exaptation. But are interlocking canines needed to fulfill this function? Well, Kiltie (1981) showed via mechanical analysis that the deeply-rooted interlocking canines were probably needed to counteract rearward torque, and you should see his paper for more details. But as always there is room for doubt.
As for the stresses involved, I don't know of bite strength analysis or finite element studies or anything like that done on peccaries, but such work might have been done. Kiltie (1982) did publish a study on peccary bite force in Biotropica but I haven't seen it.
Thanks, I'll check the references. It seems a dangerous way to reach the result but evolutions works with whatever is available.
Yes, as stated 'Use of the canines as bracing structures therefore looks like an exaptation: a new use for a set of structures that were previously used for something else'.
So the 'result' of having canines that function in bracing may be a by-product that peccaries have used to their advantage.
However, scientists still aren't sure if the fourth species of peccary is truly unique...in other news, I'm going to stick my hand in a peccary's mouth.
Wow...both interesting article and entertaining title. Peccaries are my favorites of the extant artiodactyls, and its finally nice to see them getting some love. Though even if the canines are used to lock the jaws, its still going to hurt like hell if they bite you. Keep up the great work.
Post a Comment
<< Home