Goodbye, my giant predatory, cursorial, flightless hoatzin
 Toxic Madagascan frogs are losing their toxicity, there is that ongoing controversy about the taxonomic status of the kouprey, and there have lately been some bizarre criticisms of Jeff Meldrum and his sasquatch research. One day I’ll build up enough courage to post about sasquatch, but not yet :) I am still planning to blog about Kimmeridge Clay dinosaurs and the elephant-killing lions of Chobe National Park (Botswana), not to mention temnospondyls. On a personal level, my life continues to go from bad to worse and I have been horribly ill over the past several days, but you don’t want to hear about that. Oh yeah, our house is being literally invaded by harlequin ladybirds (an alien species from Asia that arrived in Britain in 2004).
Toxic Madagascan frogs are losing their toxicity, there is that ongoing controversy about the taxonomic status of the kouprey, and there have lately been some bizarre criticisms of Jeff Meldrum and his sasquatch research. One day I’ll build up enough courage to post about sasquatch, but not yet :) I am still planning to blog about Kimmeridge Clay dinosaurs and the elephant-killing lions of Chobe National Park (Botswana), not to mention temnospondyls. On a personal level, my life continues to go from bad to worse and I have been horribly ill over the past several days, but you don’t want to hear about that. Oh yeah, our house is being literally invaded by harlequin ladybirds (an alien species from Asia that arrived in Britain in 2004).Anyway, as you’ll note from the accompanying image, I still plan for now to write about things that are related to, or inspired by, Chiappe & Bertelli’s recent paper on phorusrhacids. The image, depicting the controversial North American phorusrhacid Titanis, has been kindly provided by my good friend Carl Buell who has, I am very pleased to say, recently started blogging again after a very long absence.
In the previous post – a spinoff of a still earlier post about phorusrhacids – I discussed the South American landbird theory. It suggests that….
… there might be a hoatzin-cariamaen clade, probably persisting as relicts in
Support for this idea comes mostly from the similar hand morphology that some of these birds have, combined in part with the idea that they’re just about similar enough to be imagined as possible relatives. But if you’re only familiar with the view of bird classification presented in textbooks and so on, the idea that cariamaens might be close to such things as hoatzins is pretty surprising, as the former have conventionally been regarded as part of Gruiformes, the group that includes rails, cranes, trumpeters and several other groups. As mentioned in the previous post, there are substantial doubts however as to whether Gruiformes is monophyletic or not. What is the current thinking on this issue?
Dissipation of the gruiforms
 In their comprehensive and influential study of DNA hybridization, Sibley & Ahlquist (1990) supported gruiform monophyly, as did Livezey (1998) in a large study of morphological data, and Cracraft et al. (2004) on genetic data. However, other large-scale studies have found different gruiforms to occupy different positions within the neornithine tree (in the following discussion I have not aimed to be comprehensive: rather, I am most interested in those studies that included seriemas [and hence provide data on the position of cariamaens]).
In their comprehensive and influential study of DNA hybridization, Sibley & Ahlquist (1990) supported gruiform monophyly, as did Livezey (1998) in a large study of morphological data, and Cracraft et al. (2004) on genetic data. However, other large-scale studies have found different gruiforms to occupy different positions within the neornithine tree (in the following discussion I have not aimed to be comprehensive: rather, I am most interested in those studies that included seriemas [and hence provide data on the position of cariamaens]).
In a major and comprehensive study of morphological characters, Livezey & Zusi (2001) found gruiforms to be scattered about the neornithine tree. Seriemas were without close relatives and were the most basal group within Neoaves (the neognath clade that excludes waterfowl and gamebirds), bustards [see adjacent image] were on their own and near the middle of the neoavian radiation, trumpeters, cranes and limpkins grouped with hoatzins in a clade that also included flamingos, tubenosed seabirds, divers and penguins, and rails and finfoots were members of a ‘higher landbird’ clade. It should be noted that their paper is preliminary and that further studies (hopefully with better-resolved trees) will emerge from the immense amount of data that Livezey & Zusi collected. They noted in particular that the unusual positions they recovered for seriemas and other gruiforms were likely to change in future (p. 195).
In a study of osteological and soft-tissue characters, Mayr & Clarke (2003) also found gruiforms to be polyphyletic: rails, trumpeters and cranes (referred to from hereon as the ‘gruiform core’) were one of the most basal groups within Neoaves, bustards were without close relatives, and seriemas formed a clade with….. hoatzins. The seriema-hoatzin clade was closely allied with a cuckoo-turaco clade. The main characters tying seriemas, hoatzins, cuckoos and turacos together were those of the hand and the hip muscles, and they were also united in possessing distinctive recesses on the top of the pelvis. From the point of view of the South American landbird theory, Mayr & Clarke’s study is therefore significant in finding empirical character support for the monophyly of a turaco-hoatzin-seriema clade. However…
In a much-discussed study, Fain & Houde (2004) found that Neoaves consisted of two clades, Metaves and Coronaves. Their most exciting conclusion was that rampant convergence had occurred between these two parallel radiations: metavians include hoatzins, mesites and grebes, while coronavians include turacos, passerines and divers, for example (if those lists are lost on you, hoatzins are similar to touracos, mesites are similar to some passerines, and grebes are similar to divers). Different gruiforms were found to belong to both groups; mesites, kagus and sunbitterns were metavians close to owlet nightjars, grebes and sandgrouse; seriemas and bustards were coronavians without close relatives; while the gruiform core was part of a coronavian clade that included divers, cuckoos, turacos, tubenosed seabirds, storks, herons, penguins and pelicans.
Most recently, Ericson et al. (2006), in a study of molecular sequence data, also supported gruiform polyphyly. Their study is significant in that they found support for Fain & Houde’s Metaves-Coronaves division, and the gruiform groups fell into pretty similar positions, though with exceptions. Seriemas were not isolated within Coronaves (as they were in Fain & Houde’s study), but instead part of a clade that included parrots, passerines and falcons.
An approximate consensus
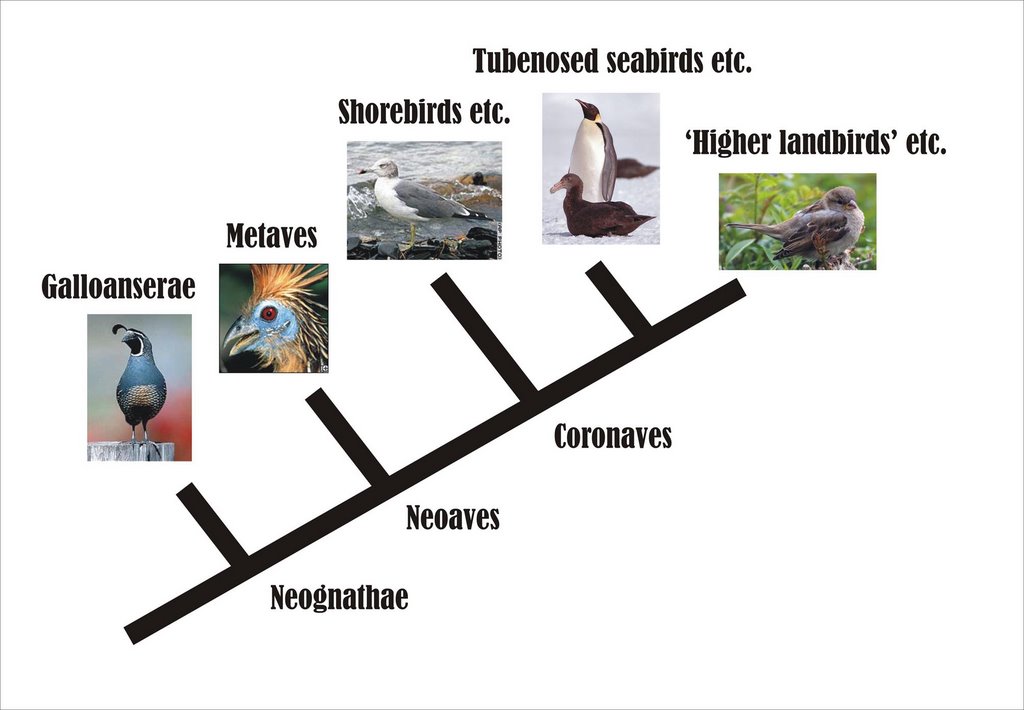
It’s difficult to conclude from these conflicting studies (and others) that we are anywhere near a consensus on neoavian affinities, but in fact we are getting somewhere, and the following details are common to all the recent major studies; gamebirds and waterfowl are at the base of Neornithes, and probably form a clade (Galloanserae) that is the sister-taxon to Neoaves; within Neoaves, tubenosed seabirds, pelicans, divers, herons and storks are part of a waterbird clade somewhere near the base of Neoaves; shorebirds (gulls, terns, skuas, auks, plovers and waders) form another clade near the base of Neoaves; and there is a ‘higher landbird’ clade that includes passerines, woodpeckers and allies, kingfishers and rollers. Owls, raptors, mousebirds, parrots, trogons, hornbills and hoopoes are most likely close to, or part of, the ‘higher landbird’ clade. Finally, there might be a hitherto overlooked metavian clade at the base of Neoaves: it includes such strange bedfellows as nightjars and other nightbirds, swifts and hummingbirds, kagus, pigeons, sandgrouse, flamingos and grebes, mesites, hoatzins and tropicbirds.
The accompanying cladogram is a highly simplified attempt at depicting this consensus.
The idea that the hoatzin is not close to seriemas or turacos, but is in fact a member of a hitherto-overlooked metavian clade at the base of Neoaves is an exciting one, mostly because it would make this bird strongly convergent on the coronavian turacos. What do other studies have to say about the affinities of the hoatzin?
The hoatzin problem
 Unfortunately the phylogenetic affinities of the hoatzin have been one of the most contested issues within avian systematics (there is an entire review article devoted to this subject: Sibley & Ahlquist 1973). Most usually considered close to either gamebirds or cuckoos (in fact, when first described in 1776 the hoatzin was classified as a species of Phasianus), hoatzins have also been allied over the years with turacos, rails, hornbills, sandgrouse and pigeons. Sibley & Ahlquist (1973) concluded that the hoatzin was not just closely related to cuckoos, but actually deeply nested within Cuculidae. This idea has been challenged by other studies, all of which find hoatzins to be outside of Cuculidae (Hughes & Baker 1999, Hughes 2000, Johnson et al. 2000), and the link with turacos has been better supported. The young of both groups clamber about among branches using their clawed fingers*, and exhibit stunted outer primaries that allow them to do this. They also share details of pterylography and soft tissue and skeletal anatomy, plus they’re generally alike in behaviour and ecology.
Unfortunately the phylogenetic affinities of the hoatzin have been one of the most contested issues within avian systematics (there is an entire review article devoted to this subject: Sibley & Ahlquist 1973). Most usually considered close to either gamebirds or cuckoos (in fact, when first described in 1776 the hoatzin was classified as a species of Phasianus), hoatzins have also been allied over the years with turacos, rails, hornbills, sandgrouse and pigeons. Sibley & Ahlquist (1973) concluded that the hoatzin was not just closely related to cuckoos, but actually deeply nested within Cuculidae. This idea has been challenged by other studies, all of which find hoatzins to be outside of Cuculidae (Hughes & Baker 1999, Hughes 2000, Johnson et al. 2000), and the link with turacos has been better supported. The young of both groups clamber about among branches using their clawed fingers*, and exhibit stunted outer primaries that allow them to do this. They also share details of pterylography and soft tissue and skeletal anatomy, plus they’re generally alike in behaviour and ecology.
* Claims that young hoatzins do not use their clawed fingers in climbing (and that they rely on the bill and feet alone) are not correct. I’ve never seen a live hoatzin, but there are many photos and bits of footage showing them climbing with their fingers.
Arguing that ‘the hoatzin problem is still unresolved’, Sorenson et al.’s (2003) new analysis of mtDNA showed that there was little or no support for the linking of hoatzins with either turacos or cuckoos, and that their data best supported an affinity between hoatzins and columbiforms (pigeons and doves). While several early avian systematists also linked hoatzins with columbiforms, Sorenson et al. (2003) noted that their results were poorly supported. Interestingly, a louse (Osculotes) unique to the hoatzin does not have any close relatives among the lice that occur on cuckoos or turacos. Of special interest to our discussion here is that Sorenson et al. (2003) didn’t include any gruiforms in their study, and hence didn’t/couldn’t test the possibility that hoatzins might be allied to any of the taxa included within that group.
Worth noting is that, while there are two fossil hoatzins, neither of them preserve enough information to tell us anything useful about hoatzin affinities, or about the way of life of the fossil forms. Hoazinoides from the Miocene of Colombia, known from a partial skull, seems to have been very similar to Opisthocomus while Onychopteryx from the Eocene of Argentina is known only from a partial tarsometatarsus and hence is not too informative.
In conclusion; Sibley & Ahlquist’s (1973) idea that hoatzins are cuckoos has now been rejected; Sorenson et al.’s (2003) conclusion that hoatzins are close to columbiforms is both poorly supported and unsatisfactory in that too few other neornithine taxa were included for comparative purposes; and studies linking hoatzins with turacos (Hughes & Baker 1999, Hughes 2000) are now questionable given that there is strong evidence from the β-fibrinogen gene that hoatzins are part of a metavian clade that does not include turacos (Fain & Houde 2004, Ericson et al. 2006).
Goodbye, my giant predatory, cursorial, flightless hoatzin
After all this then, how seriously should we take the idea that the South American landbird group is real? While recent phylogenetic studies strongly indicate that gruiforms are not monophyletic, only one large-scale study (Mayr & Clarke 2003) has found support for a clade that corresponds roughly with the South American landbird group. More recent studies, with larger data sets, have failed to group any of these birds however, and in fact hoatzins and seriemas seem to be at different ends of the neornithine tree.
In the most recent word on the subject, Ericson et al. (2006) found molecular sequence data to support a grouping of seriemas within a clade that included parrots, passerines and falcons. Is this where phorusrhacids and their relatives will finally go then? As always, we await future work, but if this view is valid, then seriemas, phorusrhacids and other cariamaens most likely evolved from small, arboreal coronavians. Their terrestrial, cursorial adaptations would then be late-evolved novelties, and not primitive features inherited from earlier neornithines.

As with any idea in science, it’s possible of course that future investigation or discovery will provide new data that supports the idea that cariamaens, hoatzins and so on are all close relatives. But for now we can reject it as poorly supported and far less well supported than other views on neornithine phylogeny. Like so many alternative theories, the South American landbird theory hinges on just a few characters that are swamped by a larger number of characters that convey a different signal. So, as appealing as it might be to imagine that condors, caracaras, turacos and hoatzins are all close kin of phorusrhacids, it really is all vague and sadly lacking in any sort of good character support. Oh well.
PS - as I write I am half-watching episode II of series 2 of the BBC’s Planet Earth. While looking at the wildlife of the Tibetan Plateau, they just featured Tibetan groundpeckers Pseudopodoces humilis, and they also just featured a Lesser florican Sypheotides indica, the bustard species pictured above.
Refs - -
Cracraft, J., Barker, F. K., Braun, M., Harshman, J., Dyke, G. J., Feinstein, J., Stanley, S., Cibois, A., Schikler, P., Beresford, P., García-Moreno, J., Sorenson, M. D., Yuri, T. & Mindell, D. P. 2004. Phylogenetic relationships among modern birds (Neornithes): towards an avian tree of life. In Cracraft, J. and Donoghue, M. (eds), Assembling the Tree of Life, pp. 468-489.
Ericson, P. G. P., Anderson, C. L., Britton, T., Elzanowski, A.,
Fain, M. G. & Houde, P. 2004. Parallel radiations in the primary clades of birds. Evolution 58, 2558-2573.
Hughes, J. M. 2000. Monophyly and phylogeny of cuckoos (Aves, Cuculidae) inferred from osteological characters. Zoological Journal of the Linnean Society 130, 263-307.
- . & Baker. Phylogenetic relationships of the enigmatic hoatzin (Opisthocomus hoazin) resolved using mitochondrial and nuclear gene sequences. Molecular and Biological Evolution 16, 1300-1307.
Johnson, K. P., Goodman, S. M. & Lanyon, S. M. 2000. A phylogenetic study of the Malagasy couas with insights into cuckoo relationships. Molecular Phylogenetics and Evolution 14, 436-444.
Livezey, B. C. 1998. A phylogenetic analysis of the Gruiformes (Aves) based on morphological characters, with an emphasis on the rails (Rallidae). Philosophical Transactions of the Royal Society of
- . & Zusi, R. L. 2001. Higher-order phylogenetics of modern Aves based on comparative anatomy.
Mayr, G. & Clarke, J. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19, 527-553.
Sibley, C. G. & Ahlquist, J. E. 1990. Phylogeny and Classification of Birds: A Study in Molecular Evolution.
- . & Ahlquist, J. E. 1973. The relationships of the hoatzin. The Auk 90, 1-13.
Sorenson, M. D., Oneal, E., García-Moreno, J. & Mindell, D. P. 2003. More taxa, more characters: the hoatzin problem is still unresolved. Molecular Biology and Evolution 20, 1484-1499.
Labels: birds, phorusrhacids
posted by Darren Naish at 8:25 PM
![]()
![]()


8 Comments:
Bird phylogenies always make my head hurt. For some reason this post
reminded me of your old post on rabbits. On the subject of the hand claws,
I remembered a post from Carel's blog where he describes making a poor
hatchling hoatzin jump into the water, but after checking it again I see
he did not take photos of the unfortunate creature climbing back.
Even if the hoatzin is not one of them, cuckoos are quite interesting.
I've seen a recent study based on osteology "behavior and ecology"
which seems to agree with molecular studies on the paraphyletic and basal
nature of terrestrial cuckoos, that arboreal cuckoos are derived
and monophyletic, and that brood parasitism evolved twice in Cuculiformes.
The paper is free but it is in Portuguese, although there's a short english
abstract and a cladogram.
Sérgio R. Posso & Reginaldo J. Donatelli (2006)
Phylogenetic analysis of Cuculiformes (Aves) based on osteology,
behavior and ecology: systematics and evolutionary implications.
Rev. Bras. Zool. v.23 n.3 Curitiba sep. 2006
Link
"I am still planning to blog about Kimmeridge Clay dinosaurs and the elephant-killing lions of Chobe National Park (Botswana), not to mention temnospondyls."
Don't tell me you've forgotten about your intention to blog on the rhinogradentia?
Viva Snouters!
Of course, if we're willing to restructure sereima relationships based on molecular analyses, who's to say phorusrhacids should follow? Isn't Cariamae itself based on the same kinds of characters that apparently incorrectly unite seriemas with hoatzins, cuckoos and turacos?
Not that I disagree with Metaves/Coronaves (quite the contrary!), and obviously we have no better way of resolving most extinct species' relationships, but that worry always crops up in my mind when I think too much about phylogeny.
Has Cariamae ever been tested in a cladistic analysis?
Thanks Mickey.. dammit, I KNEW someone would bring this up! In fact, in an earlier version of the post I did have a section where I pointed out that I am essentially assuming monophyly of Cariamae, and that it is not unquestionable. Having said that, from the morphological point of view cariamaens certainly look enough alike to be regarded as close: some phorusrhacids have previously been classified as cariamids, for example. But then does this mean anything?
Yes, Cariamae has been tested in a cladistic analysis. See....
Mayr, G. 2002. A new specimen of Salmila robusta (Aves: Gruiformes: Salmilidae n. fam.) from the Middle Eocene of Messel. Paläontologische Zeitschrift 76, 305-316.
Do so many ornithologists regard Sibley & Ahlquist 1990 as the starting point, or why do you mention it so often? After all, it's DNA-DNA hybridisation -- pure, absolute phenetics.
Interesting that you should mention Pseudopodoces since it is a very good example of how that "rampant convergence" probably starts. The groundpecker is actually a tit, more specifically it is closely related to the Great Tit, but a few million years down the road it may well end up more like a bustard or a crane.
Many other birds demonstrate the same ability to go off on an evolutionary tangent. Consider e. g. the Puna Flicker, a burrowing woodpecker who never sees a tree in his life (I barely believed my eyes when I first saw one bobbing along over the treeless puna). Or the Woodcock, a non-wading, forest-living, earthworm-eating wader. Or the Palm-nut Vulture, a near-vegetarian Accipitrid. Or the Northern Hawk-owl, a largely diurnal owl that is both behaviorally and morphologically convergent with Accipitrids.
Ah, but Mayr (2002) found that psophiids were the living sister taxon to cariamaens, correct (I don't have the paper in question)? Whereas Ericson et al. (2006) had psophiids far away in Gruiformes sensu stricto with gruids and aramids. So if the basic topology of Mayr is wrong, particular nodes could easily be wrong too. After all, topology is affected by character distribution of outgroups. Maybe things would work out differently if Mayr included falconids, psittaciformes and passeriformes. Or maybe not. I'm just playing Devil's advocate.
Thanks to all for latest comments. David: yes, Sibley & Ahlquist (1990) is often the first place people go to check ideas on neornithine phylogeny. Why? (1) Their coverage is particularly complete, with most extant genera (many not covered by any other studies) included. (2) They don't just provide their hybridisation data, but also review all previous work. Sure, it's pure phenetics, and of course the methodology and assumptions have been widely criticised, but their discoveries cannot simply be disregarded.
Tommy: yup, I specifically mentioned Pseudopodoces because of its incredible convergent similarity with corvids (whereas it now seems to actually be deeply nested within Paridae, as you note). I blogged about this species before. Go to...
http://darrennaish.blogspot.com/2006/05/more-beautifully-interesting-birds.html
Mickey: yes, you're right about Mayr's study and its limitations (I mentioned it to show that there is at least one study providing character support for cariamaen monophyly), and yes we still need bigger data sets with more taxa (including fossil ones). Livezey & Zusi's data set is shaping up this way but there is no indication that they are about to publish more papers based on their data (http://www.carnegiemnh.org/birds/porphyri.html). I still feel it's 'safe' to assume cariamaen monophyly for reasons stated previously, but, yes, we need more work.
Post a Comment
<< Home