The war on parasites: an oviraptorosaur’s eye view

In the previous post we saw how avian bill morphology is crucial to the control of parasites, an assertion that comes from recent studies of bill shape and ectoparasite control in scrub-jays, pigeons and numerous Neotropical bird species. So important is parasite control in a bird’s overall fitness that preening might be one of the most important functions of the bill and, accordingly, adaptive radiations of beak morphology should be re-assessed with both feeding and preening in mind.
The several studies that have looked at bird bills and their function in preening only considered what implications there might be for living birds. But as a palaeontologist I’m going to do the logical thing and wonder what this might mean for fossil feathered taxa. I’ll note now that the previous post is essential reading before you embark. Ok, here we go.
Fossil birds belonging to the same groups as extant species, surely, used their bills in the same manner as extant species, so dodos, teratorns and presbyornithids almost certainly found their bills to be as essential for preening as do modern pigeons, raptors and ducks. But of course we know that feathers weren’t unique to ‘modern-type’ birds: they were also present in the basal birds of the Mesozoic (going all the way back to the archaeopterygids, and including a diverse aviary of yandangornithids, confuciusornithids, enantiornitheans, hesperornitheans and others) AND they were also present in non-avian maniraptoran theropods. We know that true feathers were present in oviraptorosaurs, microraptorians (which might be part of Dromaeosauridae: it depends on the phylogeny) and almost certainly troodontids (Jinfengopteryx, a luxuriantly feathered little theropod described last year as an archaeopterygid, is almost certainly a troodontid). Furthermore, probable ‘proto-feathers’ (rather simple quill-like integumentary structures, almost certainly the morphological ancestors of true, complex feathers) were present in compsognathids, basal tyrannosauroids and alvarezsaurids.
We also know that ectoparasites were infesting feathers by the Cretaceous at least. How do we know this? Martill & Davis (1998, 2001) described an isolated feather from the Lower Cretaceous Crato Formation of Brazil that is covered in more than 240 hollow spheres that are almost certainly feather mite eggs. We also know that fleas were present in the Lower Cretaceous as there are two particularly good ones known from Australia (Riek 1970), and we also know of possible fleas and odd long-legged possibly parasitic insects from the Lower Cretaceous of the former USSR (Ponomarenko 1976). Terrestrial birds whose plumage is superficially similar to that of fuzzy small theropods are notorious for harbouring ectoparasites, with kiwis in particular being reported to crawl with numerous fleas, ticks, feather mites and lice (Kleinpaste 1991). So I would be confident that Mesozoic birds, and fuzzy and feathered non-avian theropods, had to contend with ectoparasites. What then did they do about parasite control?
Unfortunately we don’t know enough about the rhamphothecae of Mesozoic birds and bird-like maniraptorans to determine whether or not they had a maxillary overhang: the preservation simply isn’t good enough. But maybe some of these animals didn’t need a maxillary overhang given that many of them had teeth. Indeed several Mesozoic maniraptorans possess just a few teeth at the jaw tips, or even just in the upper jaw tips.
Take the feathered turkey-sized short-skulled basal oviraptorosaurs Protarchaeopteryx* and Caudipteryx (if you’ve heard that these animals aren’t oviraptorosaurs but actually flightless birds, ignore it: it’s a theory based on wishful thinking and misinterpretation of morphological evidence). In Protarchaeopteryx, teeth are restricted to the premaxillae and anterior parts of the maxillae and dentaries, with the premaxillary teeth being a few times taller than the others (Ji et al. 1998). In Caudipteryx, four procumbent teeth are present in each premaxilla, but the rest of the skull is edentulous. Incisivosaurus – closely related to, and possibly congeneric with, Protarchaeopteryx – has a reduced compliment of teeth, all of which are restricted to the anterior parts of the jaws, and two enlarged, bunny-like incisiform teeth project from each premaxilla (image at top: widely available on the web). More derived oviraptorosaurs were toothless, but the bony premaxillary margins of their upper jaw were serrated, raising the possibility that the tomium was serrated too. Could these serrations have been used in ectoparasite control? (incidentally, for more on oviraptorosaurs go to Luis Rey and the new oviraptorosaur panoply).
* Not a typo! I’ve lost count of how many times I’ve seen this name ‘corrected’ by well-meaning editors.
Among other non-avian feathered maniraptorans, it’s worth noting that microraptorians also exhibit an unusual premaxillary dentition. In Sinornithosaurus, a diastema separates the premaxillary teeth from the maxillary teeth, and the premaxillary teeth appear notably shorter than the maxillary ones (Xu & Wu 2001). While proportionally small premaxillary teeth are seen elsewhere in theropods (e.g., in tyrannosauroids), the combination of reduced dentition and diastema isn’t, and we know without question that microraptorians had complex, vaned feathers on their limbs and tails. It’s at least suggestive that the premaxillary teeth were used for preening.
Having mentioned tyrannosauroids, I might also note that basal forms combine proportionally small premaxillary teeth with quill-like integumentary structures that would have needed preening (or is grooming the correct term here?). Could those little premaxillary teeth have been specialized for ectoparasite control? I know this is grotesque speculation of the worst kind, but read on.
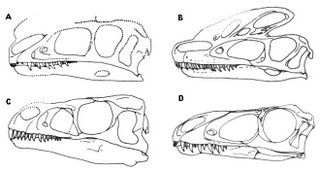
Basal tyrannosauroids and other basal coelurosaurs that possess specialised premaxillary teeth that might have been used in preening/grooming of quill-like integumentary structures. A, Proceratosaurus; B, Guanlong; C, Ornitholestes; D, Dilong. This excellent image is taken from Frederik Spindler & Peter Tschernay’s Dinosauromorpha site.
Moving now to Mesozoic birds, given that there was a trend in some lineages toward reduction and loss of teeth, it follows that members of these lineages exhibit reduced numbers of teeth relative to archaeopterygids and non-avian theropods. It seems that these birds lost the teeth from the back of the jaws first, and kept their premaxillary and dentary-tip teeth the longest. Even in forms that don’t have a reduced dentition however, we see slight heterodonty, and thus some suggestion that the anterior-most teeth were being used for something special. In archaeopterygids for example, the premaxillary teeth are more peg-like and more procumbent than are the other teeth. Aberratiodontus - an odd enantiornithean from the Chinese Jiufutang Formation - has teeth lining both its upper and lower jaws, but is reported to have rather small teeth at the jaw tips (Gong et al. 2004).
When we start looking at some of the more unusual Mesozoic birds groups, we see marked specialisation of the rostral-most dentition. Bizarre long-tailed, robust-jawed Jeholornis (almost certainly synonymous with Shenzhouraptor, and perhaps with Jixiangornis too), known from stomach contents to have eaten seeds at least occasionally, has just three very small teeth at each lower jaw tip (Zhou & Zhang 2002): the upper jaw was edentulous.
The unusual long-armed Sapeornis, also from the Jiufutang Formation, had a rather short, Caudipteryx-like skull, and short, conical, unserrated, procumbent teeth projected from its premaxillae (Zhou & Zhang 2003). Its dentaries were toothless (and its maxillae probably were too). The somewhat similar Omnivoropteryx, also from the Jiufutang Formation, was also short-skulled, and also has just a few procumbent teeth restricted to the premaxillae (Czerkas & Ji 2002). Most (but not all) enantiornitheans were toothed, and ancestrally they had teeth lining their upper and lower jaws as archaeopterygids did. But in the Yixian Formation enantiornithean Protopteryx (image at left: taken from Dinosaur.net.cn) there are just two conical, unserrated teeth in the premaxillae and two subtriangular teeth at the dentary tips (Zhang & Zhou 2000). It doesn’t seem that having a total of four teeth is a tremendously useful thing if you need those teeth to procure or dismember your food, and it’s intriguing that Protopteryx sports highly elongate, strap-like rectrices that would (presumably) have needed careful preening. The euenantiornithean Eoenantiornis has four subconical teeth in each premaxilla while there were probably six or seven teeth in each dentary, the rostral-most two of which were larger than the others (Zhou et al. 2005). Long-skulled Longirostravis has ten small, conical teeth restricted entirely to its slim jaw tips (Hou et al. 2004) and short, conical teeth are similarly only at the jaw tips in another long-skulled enantiornithean, Longipteryx (Zhang et al. 2001). And there are yet other examples of this sort of thing.
Most (but not all) enantiornitheans were toothed, and ancestrally they had teeth lining their upper and lower jaws as archaeopterygids did. But in the Yixian Formation enantiornithean Protopteryx (image at left: taken from Dinosaur.net.cn) there are just two conical, unserrated teeth in the premaxillae and two subtriangular teeth at the dentary tips (Zhang & Zhou 2000). It doesn’t seem that having a total of four teeth is a tremendously useful thing if you need those teeth to procure or dismember your food, and it’s intriguing that Protopteryx sports highly elongate, strap-like rectrices that would (presumably) have needed careful preening. The euenantiornithean Eoenantiornis has four subconical teeth in each premaxilla while there were probably six or seven teeth in each dentary, the rostral-most two of which were larger than the others (Zhou et al. 2005). Long-skulled Longirostravis has ten small, conical teeth restricted entirely to its slim jaw tips (Hou et al. 2004) and short, conical teeth are similarly only at the jaw tips in another long-skulled enantiornithean, Longipteryx (Zhang et al. 2001). And there are yet other examples of this sort of thing.
So far as I can see from all these unusual patterns of dentition, there are three possible explanations:-
(1) as dental reduction occured, a gradual step-wise loss of teeth simply meant that premaxillary and/or dentary-tip teeth were the last to go;
(2) premaxillary and/or dentary-tip teeth were retained in specialized taxa that used those teeth to procure or dismember whatever it was that they were eating;
(3) premaxillary and/or dentary-tip teeth were retained - even when not essential to foraging or feeding - as they were used in ectoparasite control.
While it’s nice to speculate – and so far that’s all I’ve done here – how might we test the idea that these Mesozoic taxa were using their unusual rostral teeth to preen with? Herein lies the rub, as I can't think of a reliable test. So far as we know, feathers aren’t abrasive enough to leave any sort of distinctive microwear on teeth, or even on rhamphothecae, so there isn’t going to be any sort of tooth wear that can be correlated with preening. It’s possible that there might be some sort of correlation between tooth spacing and feather morphology, but I find this unlikely.
Conversely, we can test the idea that teeth were used in feeding, as feeding does leave visible sorts of micro- or macrowear. Earlier I mentioned the bunny-like teeth of Incisivosaurus, and because its incisiform teeth do exhibit wear facets, they were almost certainly used in feeding. This confirms ‘explanation 2’ given above, and therefore indicates that ‘explanation 3’ didn’t apply in this case. But the two ‘explanations’ aren’t mutually exclusive, as the teeth could still have been important in ectoparasite control.
Those short premaxillary teeth present in tyrannosauroids have conventionally been regarded as having a primary role in feeding, and it might be easy to confirm this by looking for micro- or macrowear. And yes, I consider it highly highly speculative to wonder if those teeth might have functioned in grooming/preening, but I couldn’t resist mentioning it (I want to discuss here the SEM data I have on the premaxillary teeth of the basal tyrannosauroid Eotyrannus, but that would require adding too many extra words).
Finally, if specialised teeth could be shown to have no important function in foraging or feeding behaviour it might then be logical to infer that preening was their primary function - - but, how on earth would you show that they had ‘no function’ in foraging or feeding behaviour? This just isn’t possible in Mesozoic animals when so little is known of their ecology. An analogy does come to mind: it’s been shown that the unusual dentary teeth of Impala Aepyceros melampus have a morphology specialised for a primary function in grooming. If impalas were extinct I suppose it’s possible that people might have worked this out, but how would you verify it?
Does anyone have any better ideas?
I’m far from the first person to look at Mesozoic feathered theropods this way – many other people have mentioned these ideas before, and artists have even illustrated ectoparasite control in dinosaurs. In Dinosaurs of the Air Greg Paul illustrated a Sinosauropteryx scratching in order to remove ectoparasites, and the cover of The Dinosauria, Second Edition (the current industry-standard volume on dinosaurs) features a Sinosauropteryx (this time by Mark Hallett) nibbling at its proto-feathers, again presumably as a form of ectoparasite control.
But I don’t think anyone’s really married data on Mesozoic birds and other theropods with the new work of Dale Clayton and colleagues on ectoparasite control in extant birds. Maybe this idea will bear proverbial fruit down the line, but for now this is where my contribution ends.
Finally, here’s another spin on this subject. Theropods weren’t the only Mesozoic tetrapods with a furry coat of integumentary fibres: we also know that pterosaurs were fuzzy too. So did they also have to contend with ectoparasites? I’ll say no more on this topic, but perhaps it can be elaborated on at another time. For the latest news on Tetrapod Zoology do go here.
Refs - -
Czerkas, S. A. & Ji, Q. 2002. A preliminary report on an omnivorous volant bird from northeast China. In Czerkas, S. J. (ed) Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum (Blanding, Utah), pp. 127-135.
Gong, E., Hou, L. & Wang, L. 2004. Enantiornithine bird with diapsidian skull and its dental development in the Early Cretaceous in Liaoning, China. Acta Geologica Sinica 78, 1-7.
Hou, L., Chiappe, L. M., Zhang, F. & Chuong, C.-M. 2004. New Early Cretaceous fossil from China documents a novel trophic specialization for Mesozoic birds. Naturwissenschaften 91, 22-25.
Ji, Q., Currie, P. J., Norell, M. A. & Ji, S. 1998. Two feathered dinosaurs from northeastern China. Nature 393, 753-761.
Kleinpaste, R. 1991. Kiwis in a pine forest habitat. In Fuller, E. (ed) Kiwis: A Monograph of the Family Apterygidae. Swan Hill Press (Shrewsbury), pp. 97-138.
Martill, D. M. & Davis, P. G. 1998. Did dinosaurs come up to scratch? Nature 396, 528-529.
- . & Davis, P. G. 2001. A feather with possible ectoparasite eggs from the Crato Formation (Lower Cretaceous, Aptian) of Brazil. Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen 219, 241-259.
Ponomarenko, A. G. 1976. A new insect from the Cretaceous of Transbaikalia, a possible parasite of pterosaurians. Paleontology Journal 1976 (3), 339-43.
Riek, E. F. 1970. Lower Cretaceous fleas. Nature 227, 746-747.
Xu, X. & Wu, X.-C. 2001. Cranial morphology of Sinornithosaurus millenii Xu et al. 1999 (Dinosauria: Theropoda: Dromaeosauridae) from the Yixian Formation of Liaoning, China. Canadian Journal of Earth Sciences 38, 1739-1752.
Zhang, F. & Zhou, Z. 2000. A primitive enantiornithine bird and the origin of feathers. Science 290, 1955-1959.
- ., Zhou, Z., Hou, L. & Gu, G. 2001. Early diversification of birds: evidence from a new opposite bird. Chinese Science Bulletin 46, 945-949.
Zhou, Z., Chiappe, L. M. & Zhang, F. 2005. Anatomy of the Early Cretaceous bird Eoenantiornis buhleri (Aves: Enantiornithes) from China. Canadian Journal of Earth Sciences 42, 1331-1338.
- . & Zhang, F. 2002. A long-tailed, seed-eating bird from the Early Cretaceous of China. Nature 418, 405-409.
- . & Zhang, F. 2003. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. Journal of Paleontology 40, 731-747.
posted by Darren Naish at 9:40 PM
![]()
![]()


7 Comments:
Cool stuff! I was wondering, have feathered dinosaurs been found in places other than China? (And if not, do you think it's a) because the sediment in Liaoning was fine and good for preserving feathers, or b) because feathered dinosaurs just weren't that common in other parts of the world? (Sorry for that incredibly stilted question, haha...)
Hi, thanks for the comments. To date, non-avian theropods that have feathers preserved are unique to a few sedimentary units (including the Qiaotou, Jiufotang and Yixian formations) in China.
This certainly results from the fact that this is the only place in the world where we have small theropods preserved in so-called Lagerstatten deposits (that is, places where exceptionally good preservation of soft tissues occurred). In other words, it's a sampling fluke.
What about ornithischians, Effigia, and turtles? They all have beaks, yet no feathers to preen. When talking about non-avian theropods, you have to be careful about assigning a primary function to a presumably multifunctional structure, that may or may not have intially evolved for its current use.
I'd appreciate it if people didn't leave unattributed comments. Please sign off with your name.
Regardless... there is nothing is this post which equates to 'rhamphothecae evolved for parasite control'. It is clear (I made the point a few times in the previous post) that ectoparasite control is one of the bill's functions, but it does not follow that bills evolved for ectoparasite control. So the presence of beaks in unfeathered taxa is not a problem. And anyway, it's not the presence of a beak that's important: it's specialisations of the beak tissue, such as the maxillary overhangs.
In other words, I am not saying that parasite control is a primary function of beaks and unusual teeth: rather, I was wondering if these structures might have functioned in this role.
As a guess, does your surname being with the letter 'I'?
Protarchaeopteryx is not just not a typo, it's even etymologically correct. Greek and Latin don't like vowel clusters! (Confuciusornis, on the other hand, hurts.)
Protopteryx is not from the Yixian Fm but from the Dabeigou Fm (Hebei, not Liaoning) which is supposed to be a bit older.
How actually did you manage to count teeth in Protarchaeopteryx? The photos I've seen are coarse-grained, and the fossil itself seems to have been split into two slabs as usual.
Also, what about Hesperornithes and Ichthyornis which have teeth everywhere except the pmx?
Re: Protopteryx, in my defence I should point out that Zhang & Zhou (2000) did initially report the taxon as coming from the Yixian Formation, and that the distinction of the Yixian and the Dabeigou has been controversial: He et al. have a paper about this in the latest Geochemistry, Geophysics, Geosystems. Has the distinction been widely accepted formally?
How did I manage to count the teeth in Protarchaeopteryx? I didn't! There's nothing in the text on a tooth count for this taxon. I will note here that I've closely examined the holotype more than once.
Finally, yes there are toothed birds in which the distribution of teeth does not appear to make sense for a preening function. Well then: these birds did not use their teeth in preening! I am NOT saying that yandangornithids and so on have teeth for preening, I'm wondering if the teeth might have functioned in this role.
"Re: Protopteryx, in my defence I should point out that Zhang & Zhou (2000) did initially report the taxon as coming from the Yixian Formation, and that the distinction of the Yixian and the Dabeigou has been controversial: He et al. have a paper about this in the latest Geochemistry, Geophysics, Geosystems. Has the distinction been widely accepted formally?"
No idea, I've found the term "Dabeigou Fm" in one paper (description of Hongshanornis) and "Qiaotou Fm" in one other (description of Jinfengopteryx). Thanks a lot for alerting me to that paper, I wouldn't have found it otherwise!!! (I have stumbled across the one by He et al. on the age of the Daohugou bed, though -- Valanginian or younger, so maybe Barremian again.)
Post a Comment
<< Home