We flightless primates
 Mention ‘flying primate’ and most zoologists will think you’re referring to the well known, controversial theory of John Pettigrew of the University of Queensland. Initially basing his theory on retinotectal organization (viz, the way in which data from the retina is processed in the brain), Pettigrew (1986) argued that megabats (the group that includes fruit bats) are not close relatives of microbats (the mostly small, mostly insectivorous bats that mostly use echolocation), but that they’re actually flying primates, of a sort (read on). If this is right, then Chiroptera [the proper name for the bat clade] is not monophyletic, and true flight evolved at least twice among mammals. Furthermore, Primates and their close relatives are more diverse than conventionally thought, and include an impressive radiation of hitherto-misclassified volant species.
Mention ‘flying primate’ and most zoologists will think you’re referring to the well known, controversial theory of John Pettigrew of the University of Queensland. Initially basing his theory on retinotectal organization (viz, the way in which data from the retina is processed in the brain), Pettigrew (1986) argued that megabats (the group that includes fruit bats) are not close relatives of microbats (the mostly small, mostly insectivorous bats that mostly use echolocation), but that they’re actually flying primates, of a sort (read on). If this is right, then Chiroptera [the proper name for the bat clade] is not monophyletic, and true flight evolved at least twice among mammals. Furthermore, Primates and their close relatives are more diverse than conventionally thought, and include an impressive radiation of hitherto-misclassified volant species.In more detailed papers, Pettigrew (1991) and Pettigrew et al. (1989) marshaled evidence from eye, brain and spinal cord anatomy, fore- and hindlimb, finger and metacarpal proportions, and haemoglobin sequences, and again concluded that megabats and primates shared a common ancestor, and that microbats were not close relatives of megabats, but that their affinities lay elsewhere. Pettigrew et al. (1989) further argued that colugos (aka flying lemurs, or dermopterans) were also part of the megabat-primate clade, and essentially late-surviving relics which resembled the common ancestor of the megabat-primate clade. Pettigrew and colleagues weren’t the first to question bat monophyly: John E. Hill of the then British Museum (Natural History) had done this as early as 1976, Smith & Madkour (1980) argued that micro- and megabats were of separate origins, and Hill & Smith (1984), in one of the best and oft-cited overviews on bat evolution and biology, expressed scepticism of bat monophyly and a preference for megabat-primate affinities (p. 36).
Popularly, Pettigrew et al.’s notion that megabats are closer to primates than to microbats became characterized as the ‘flying primate’ theory, and Pettigrew et al. used this term in their papers. This characterization isn’t accurate however as Pettigrew et al. (1989) specifically stated that, within their favoured phylogenetic scheme, megabats would be outside of the clade Primates, and thus not primates in the true sense (p. 551).

I must admit that the ‘flying primate’ theory has a great deal of intuitive appeal, and this probably explains why it’s become both well known and much written about. Colin Tudge wrote about it in The Independent for example (his article was titled ‘That’s no bat, that’s my brother'), and it isn’t every day that problem areas within the higher-level phylogenetics of placental mammals make it into daily newspapers. Why is the ‘flying primate’ hypothesis intuitively appealing? Perhaps because it appears to have reasonable and easily understood character support and is exciting in contending that a complex and strongly modified bit of morphology – namely the bat wing, with its bizarre elongated fingers and patagial membranes – evolved independently more than once.
The accompanying image by Peter Schouten [click for larger version], commissioned by John Pettigrew, depicts the 'flying primate' hypothesis in graphic form: note that colugos and megabats branch off from the primate lineage, and aren't alongside microbats. This image is borrowed from Pettigrew's Neuroscience UQ site.
However, from the start most bat experts had a problem with the concept: Wible & Novacek (1988) showed how numerous skeletal and soft tissue features ‘strongly support the inclusion of megabats and microchiropterans within the single order Chiroptera’ (p. 1). Simmons et al. (1991) argued that Pettigrew’s entire approach to the bat monophyly question was flawed, Thewissen & Babcock (1992) argued that the detailed anatomy of wing musculature best supported bat monophyly, while Bailey et al. (1992) and Ammermann & Hillis (1992) found that genetic data better supported the case for bat monophyly than that for diphyly. Bailey et al.’s paper is even titled ‘Rejection of the ‘flying primate’ hypothesis’, and in an accompanying news piece in Science Ann Gibbons wrote how the ‘flying primate’ hypothesis was ‘heading for a crash landing’. A comprehensive overview of all the data supporting bat monophyly was provided by Nancy Simmons (1994).
However, DNA-based studies later produced by Pettigrew and his colleagues produced a new, even more surprising result: rhinolophoids (the horseshoe bats and their relatives) were consistently found to group together with megabats, the implication now being that Microchiroptera might be non-monophyletic (Hutcheon et al. 1998, Kirsch & Pettigrew 1998, Pettigrew & Kirsch 1998). The existence of a rhinolophoid-megabat clade has also been supported by other research teams (e.g., Teeling et al. 2000, 2002, Liu et al. 2001) but note that many of these studies (Hutcheon et al. 1998, Teeling et al. 2000, 2002, Liu et al. 2001) did not support Pettigrew's idea of chiropteran diphyly.
Let’s suppose for a moment however that Pettigrew and colleagues are right, and that neither Chiroptera nor Microchiroptera are monophyletic. Suppose also that bats are members of the placental mammal group Archonta – an idea that was quite widely accepted until recently (read on). Archonta was first named by William King Gregory in 1910* to house Scandentia (tree shrews), Chiroptera, Dermoptera and Primates (Gregory also included sengis/elephant shrews within Archonta, but they were later removed from it as their affinities clearly lie elsewhere). As the only mammalian clade whose constituent groups are all tied to a climbing lifestyle, it was always very nice to have the bats in there, given that an arboreal origin for bat flight has always been favoured by everyone who’s ever pondered the issue.
* It’s always been problematical that archontan monophyly has only ever been supported by two morphological characters: the anatomy of the penis, and the shape of one of the facets on the astragalus. By their genitals and ankles ye shall know them. If you’re wondering, Archonta means ‘ruling beings’. Yuck.
As you see from the little cladogram I’ve knocked up here [click for larger version], bat diphyly and archontan monophyly makes it at least possible – and phylogenetically parsimonious – that flight was primitive for the megabat-primate clade, or in other words that primates are secondarily flightless. That’s right: you and me, and other apes, and lemurs, tarsiers and monkeys all descend from flying, winged proto-primates. If I were George Olshevsky I might dub this the ‘bats come first’ theory.
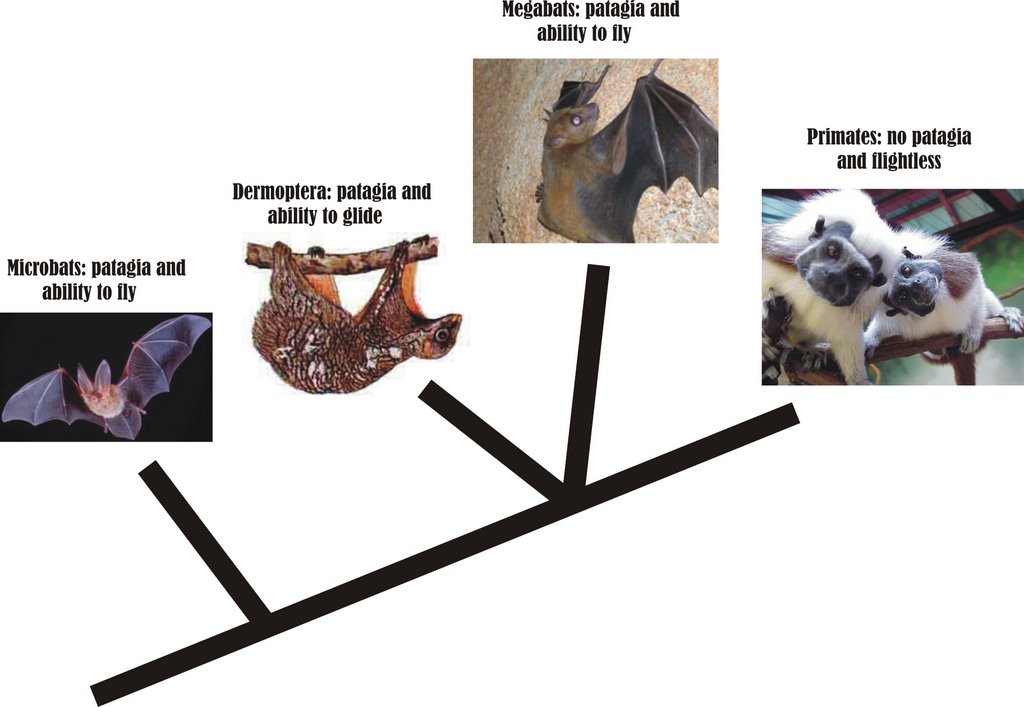
But, no, it wasn’t to be. Bat diphyly is not currently favoured by the evidence, and even Archonta has now fallen by the wayside: DNA-based phylogenies now indicate that, while Scandentia, Dermoptera and Primates probably do form a clade – the newly dubbed Euarchonta – bats aren’t close relatives of euarchontans at all. Instead they go elsewhere within the placental mammal clade Laurasiatheria, being (most surprisingly) closest to carnivorans, perissodactyls and artiodactyls (e.g., Murphy et al. 2001, Liu et al. 2001). And what of the idea that microbats aren’t monophyletic? Alas, a recent review found that this arrangement was not as well supported by the data as is the traditional view of microbat monophyly (Jones et al. 2002).
As always the full story that I wanted to cover is even more complex than what’s related here. I haven’t even mentioned the paromomyids: a group of Eocene euarchontans proposed at one time to be colugo-like gliders and thus implicated in the issue of primate and colugo origins. Nor have I discussed the little nyctitheres, a poorly known and long-mysterious Eocene-Oligocene group that appear to have been primitive, scansorial relatives of euarchontans. In fact comparison of nyctitheres with bats and euarchontans led Hooker (2001) to restate the case for archontan monophyly, with Deccanolestes from the Late Cretaceous of India being the oldest member of the group according to his study. Genetic data clearly doesn’t favour archontan monophyly right now, but there is still at least some morphological support for it. A common problem.
Here’s the great irony of this post. I started writing about the whole ‘flying primate’ thing because I planned to use it as the introductory few paragraphs for a different topic. So while most zoologists think Pettigrew and megabats when hearing of ‘flying primates’, far less well known is that primates proper – and let me make it clear this time that I really do mean primates in the traditional sense (viz, without the megabats) – do however include species that fly. Well, fly sort of. More to come soon.
For previous posts on bats see Greater noctules: specialist predators of migrating passerines and Chewed bones and bird-eating microbats. For stuff on vampire bats please go here.
Update (added 5-8-2006): a new paper just published by Nishahara et al. (2006) has reported new genetic support for a laurasiatherian clade composed of bats, perissodactyls, carnivorans and pangolins. They name this clade Pegasoferae, a name derived by uniting Pegasus (in their view a sort of bat-perissodactyl combination) with Ferae (the name they use for the carnivoran + pangolin clade). Thanks to Stephen Bodio's Querencia and microecos.
Nishihara, H., Hasegawa, M. & Okada, N. 2006. Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions. Proceedings of the National Academy of Sciences 103, 9929-9934. Pdf here.
Refs - -
Ammerman, L. K. & Hillis, D. M. 1992. A molecular test of bat relationships: monophyly or diphyly? Systematic Biology 41, 222-232.
Bailey, W. J., Slightorn, J. L. & Goodman, M. 1992. Rejection of the ‘flying primate’ hypothesis by phylogenetic evidence from the globin gene. Science 256, 86-89.
Hill, J. E. & Smith, J. D. 1984. Bats: a Natural History. British Museum (Natural History), London.
Hooker, J. J. 2001. Tarsals of the extinct insectivoran family Nyctitheriidae (Mammalia): evidence for archontan relationships. Zoological Journal of the Linnean Society 132, 501-529.
Hutcheon, J. M., Kirsch, J. A. W. & Pettigrew, J. D. 1998. Base-compositional biases and the bat problem. III. The question of microchiropteran monophyly. Philosophical Transactions of the Royal Society of London B 353, 607-617
Jones, K. E., Purvis, A., MacLarnon, A., Bininda-Emonds, O. R. P. & Simmons, N. B. 2002. A phylogenetic supertree of the bats (Mammalia: Chiroptera). Biological Reviews 77, 223-259.
Kirsch, J. A. W. & Pettigrew, J. D. 1998. Base-compositional biases and the bat problem. II. DNA-hybridization trees based on AT- and GC-enriched tracers. Philosophical Transactions of the Royal Society of London B 353, 381-388.
Liu, F.-G. R., Miyamoto, M. M., Freire, N. P., Ong, P. Q., Tennant, M. R., Young, T. S. & Gugel, K. F. 2001. Molecular and morphological supertrees for eutherian (placental) mammals. Science 291, 1786-1789.
Murphy, W. J., Eizirik, E., Johnson, W. E., Zhang, Y. P., Ryder, O. A. & O’Brien, S. J. 2001. Molecular phylogenetics and the origins of placental mammals. Nature 409, 614-618.
Pettigrew, J. D. 1986. Flying primates? Megabats have the advanced pathway from eye to mid-brain. Science 231, 1304-1306.
- . 1991. Wings or brains? Convergent evolution in the origin of bats. Systematic Zoology 40, 199-216.
- . & Kirsch, J. A. W. 1998. Base-compositional biases and the bat problem. I. DNA-hybridization melting curves based on AT- and GC-enriched tracers. Philosophical Transactions of the Royal Society of London B 353, 369-370.
- ., Jamieson, B. G. M., Robson, S. K., Hall, L. S., McAnally, K. I. & Cooper, H. M. 1989. Phylogenetic relations between microbats, megabats and primates (Mammalia: Chiroptera and Primates). Philosophical Transactions of the Royal Society of London B 325, 489-559.
Simmons, N. B. 1994. The case for chiropteran monophyly. American Museum Novitates 3103, 1-54.
- ., Novacek, M. J. & Baker, R. J. 1991. Approaches, methods, and the future of the chiropteran monophyly controversy: a reply to J. D. Pettigrew. Systematic Zoology 40, 239-243.
Smith, J. D. & Madkour, G. 1980. Penial morphology and the question of chiropteran monophyly. In Wilson, D. E. & Gardner, A. L. (eds) Proceedings of the 5th International Bat Research Conference. Texas Tech Press (Lubbock), pp. 347-365.
Teeling, E. C., Madsen, O., Van Den Bussche, R. A., de Jong, W. W., Stanhope, M. J. & Springer, M. S. 2002. Microbat paraphyly and the convergent evolution of a key innovation in Old World rhinolophoid microbats. Proceedings of the National Academy of Sciences 99, 1431-1436.
- ., Scally, M., Kao, D. J., Romagnoll, M. L., Springer, M. S. & Stanhope, M. J. 2000. Molecular evidence regarding the origin of echolocation and flight in bats. Nature 403, 188-192.
Thewissen, J. G. M. & Babcock, S. K. 1992. The origin of flight in bats. BioScience 42 (5), 340-345.
Wible, J. R. & Novacek, M. J. 1988. Cranial evidence for the monophyletic origin of bats. American Museum Novitates 2911, 1-19.
posted by Darren Naish at 9:25 PM
![]()
![]()


14 Comments:
Do you happen to know where the name Euarchonta was coined?
Yup, it was first in...
Waddell, P. J., Okada, N. & Hasegawa, M. 1999. Towards resolving the interordinal relationships of placental mammals. Systematic Biology 48, 1-5.
Interesting post. I've also recently posted regarding the supraordinal clade euarchontaglires and the Asian origin of primates at Sinanthropus
Thanks a lot!
So where does this fit in?
I know sequence data isn't the be all, end all of phylogenetic analysis, but couldn't sequence data settle this controversy?
This is a fascinating post.
One minor detail I wanted to point out: "archonta" is a participle that comes from a Greek verb stem that yields many words dealing with power, yes, (archon, monarch), but also with priority, antiquity, and the past (archaeology, archaic). It could as easily mean "beginning ones" as "ruling ones." This may well be the meaning invoked in the naming of the group.
Thanks for your comment Kate.
Ok, Archonta might mean 'beginning ones', but in this case we know that Gregory specifically meant it to mean 'ruling ones'. His thinking was that we humans - as the 'dominant' forms of life on the planet - are 'ruling animals'. He may or may not have been inspired by Archosauria Cope, 1869: the 'ruling reptile' group that includes dinosaurs.
In Athens the Archontes were quite powerful men...
The link to worth1000.com doesn't work.
Sequence data? The whole post talks about sequence data all the time!
For what it's worth, the worth1000.com link was a photo composite of a chimp with bird wings.
Impressive birds, ground hornbills-- I saw many pairs walking around on the ground in Zimbabwe, near Kariba, years ago-- you could see those white- tipped black wings for a mile.
You can see a fine gallery of hornbills including a ground species in Carel's book. (Lots of other good stuff for Tetrapod Zoology fans too).
Well, many Homo sapiens from about a century ago on have flown, sort of. (I myself have [sort of] flown on a number of occasions over the past decade, most recently to and from the ISPN meeting in Yale earlier this summer.) But I have a feeling you mean something else?...
--
Mike Keesey
Pettigrew's QU Home Page has the article link ?Funeral of Flying Primate Hypothesis which makes for salutary reading on misleading DNA comparisons.
Re Pettigrew, it is the paper "Summary of Studies on Primate Phylogeny and 'Flying Primate' Hypothesis" rather than the earlier one which studies flawed DNA analysis in detail.
Post a Comment
<< Home