 These are exciting times, if you’re into uber-nerdy zoological discoveries. At long long last, New Zealand’s Miocene fossil mammals have finally been published. Not only do these fossils show that terrestrial mammals were formerly present on New Zealand, they are remarkable in apparently belonging to an animal that must have diverged during the Cretaceous. If you’re wondering: yes, this is the fossil that inspired my previous posts on late-surviving non-mammalian synapsids (here and here). And also on Mesozoic mammals we have the amazing new gliding mammal Volaticotherium [image at left] from the controversial Daohugou beds of Inner Mongolia. Other neat recent discoveries – from the Cretaceous – include the new Mongolian dromaeosaur Tsaagan mangas and the (allegedly) flightless enantiornithean Elsornis keni. I’ll blog about all of these things, time permitting (famous last words).
These are exciting times, if you’re into uber-nerdy zoological discoveries. At long long last, New Zealand’s Miocene fossil mammals have finally been published. Not only do these fossils show that terrestrial mammals were formerly present on New Zealand, they are remarkable in apparently belonging to an animal that must have diverged during the Cretaceous. If you’re wondering: yes, this is the fossil that inspired my previous posts on late-surviving non-mammalian synapsids (here and here). And also on Mesozoic mammals we have the amazing new gliding mammal Volaticotherium [image at left] from the controversial Daohugou beds of Inner Mongolia. Other neat recent discoveries – from the Cretaceous – include the new Mongolian dromaeosaur Tsaagan mangas and the (allegedly) flightless enantiornithean Elsornis keni. I’ll blog about all of these things, time permitting (famous last words). On the subject of fossils, a major personal event occurred today, and it involves SVP (the Society of Vertebrate Paleontology). To whom it concerns: thank you, sincerely. Anyway, moving on: what about those goddam seals?
 One of the most asked about questions I’ve encountered in tetrapod zoology concerns the mysterious seals of Siberia’s Lake Baikal. Everyone knows that an endemic, particularly small species of land-locked freshwater seal lives there, but nobody really knows how it got there. Less well known is that it isn’t the only lake-dwelling seal in the world: there are populations of Ringed seals Phoca hispida in Lake Saimaa (P. h. saimensis) and Lake Ladoga (P. h. ladogensis) in Fennoscandia, there is the Caspian seal P. caspica, and there are freshwater populations of the Common or Harbour seal P. vitulina in the Lacs des Loups Marins of northern Quebec, and in Alaska’s Lake Iliamna (Smith et al. 1996). Furthermore, Ringed seals became isolated in the Baltic Sea about 12,500 years ago when the connection with the North Sea closed due to glaciation, and effectively found themselves in a giant enclosed inland lake. I was planning to discuss these other forms, but have run out of space and time.
One of the most asked about questions I’ve encountered in tetrapod zoology concerns the mysterious seals of Siberia’s Lake Baikal. Everyone knows that an endemic, particularly small species of land-locked freshwater seal lives there, but nobody really knows how it got there. Less well known is that it isn’t the only lake-dwelling seal in the world: there are populations of Ringed seals Phoca hispida in Lake Saimaa (P. h. saimensis) and Lake Ladoga (P. h. ladogensis) in Fennoscandia, there is the Caspian seal P. caspica, and there are freshwater populations of the Common or Harbour seal P. vitulina in the Lacs des Loups Marins of northern Quebec, and in Alaska’s Lake Iliamna (Smith et al. 1996). Furthermore, Ringed seals became isolated in the Baltic Sea about 12,500 years ago when the connection with the North Sea closed due to glaciation, and effectively found themselves in a giant enclosed inland lake. I was planning to discuss these other forms, but have run out of space and time.
First named by Johann Friedrich Gmelin (1748-1804) in 1788*, the Baikal seal (also known as the Nerpa) was mentioned in print as early as 1763. Much exploited by local people for its meat and pelt, it was seriously reduced in numbers during the 1930s, and during the 1970s 2-3000 pups were being harvested each year for their skins. Bonner (1994) later gave higher numbers of 5-6000, implying that harvest numbers have increased within recent decades. A survey performed in 2000 revealed the presence of about 85,000 seals in total, and their numbers are reported to be falling as increasing numbers of pups are being killed.
* Incidentally, Gmelin thought that Baikal seals were just a form of Common seal, and not a distinct species.
 Outbreaks of canine distemper virus resulted in the deaths of about 5000 Baikal seals during 1987-88, and it is inferred that the seals were infected by domestic dogs at some stage (Kennedy et al. 2000). Exactly how this happened is not clear, though canine distemper has also caused die-offs in Antarctic Crabeater seals Lobodon carcinophagus and other pinnipeds. This problem has afflicted Asia’s other land-locked seal, the Caspian seal. Between April and May 2000, over 10,000 Caspian seals are estimated to have died along the coast of Kazakhstan, with high death rates also reported from the coasts of Azerbaijan and Turkmenistan. Necropsies demonstrated that canine distemper virus was also the primary cause of death for all of the dead Caspian seals.
Outbreaks of canine distemper virus resulted in the deaths of about 5000 Baikal seals during 1987-88, and it is inferred that the seals were infected by domestic dogs at some stage (Kennedy et al. 2000). Exactly how this happened is not clear, though canine distemper has also caused die-offs in Antarctic Crabeater seals Lobodon carcinophagus and other pinnipeds. This problem has afflicted Asia’s other land-locked seal, the Caspian seal. Between April and May 2000, over 10,000 Caspian seals are estimated to have died along the coast of Kazakhstan, with high death rates also reported from the coasts of Azerbaijan and Turkmenistan. Necropsies demonstrated that canine distemper virus was also the primary cause of death for all of the dead Caspian seals.
Despite its early discovery, the Baikal seal remained all but unknown to western scientists until the 20th century, and only in 1909 did specimens first arrive in Britain. These were collected by Charles Hose who was using the Trans-Siberian railway to get to Sarawak (which is where, in 1895, he discovered the cetacean that later became known as Fraser’s dolphin Lagenodelphis hosei). During a two-day stop at Lake Baikal, Hose managed to get local fishermen to catch three of the seals for him, alive, and he then resumed the train journey with the seals stuck in the luggage racks of his train compartment. Two of the seals died and Hose performed dissections on them while still in the carriage, ‘flinging the more perishable parts out of the train window, to the consternation of fellow passengers’ (King 1983, p. 92). The third seal died while on a ship bound for Shanghai. This little-known information was published in Hose’s autobiographical work of 1927, amusingly titled Fifty Years of Romance and Research or a Jungle-Wallah at Large. I’ve never seen this book, but the information is repeated in Judith King’s excellent Seals of the World. The first live Baikal seals weren’t seen in Britain until 1959 when Moscow Zoo sent a pair to London Zoo.
 Baikal seals are morphologically interesting for a number of reasons. Firstly, they are one of the smallest seals, reaching just 1.4 m and 80-90 kg at most (Ringed seals are smaller, as are members of some lake-dwelling Common seal populations). Secondly, for their size they have among the biggest eyes of any pinniped, with the eyeballs being so big that they almost contact one another along the skull midline. Thirdly, the huge eyes appear to have forced other skull structures to become compressed or reduced: the frontal sinuses are strongly compressed and located further ventrally than is normal for seals; the nasal cavity has been forced into an unusually low position; and the brain has been displaced backwards. The jaw muscles, the ear region and the bones around the jaw joint are also modified relative to the condition in other seals, mostly in being smaller and weaker (Endo et al. 1998a, b, 1999). The foreflippers and their claws are larger and stronger in Baikal seals than is usual for seals, and the foreclaws are distinctive in being triangular in cross-section and in having a marked dorsal ridge at their distal end. It is inferred that these forelimb structures make Baikal seals better at making and maintaining breathing holes and grasping prey than other seals (Thomas et al. 1982).
Baikal seals are morphologically interesting for a number of reasons. Firstly, they are one of the smallest seals, reaching just 1.4 m and 80-90 kg at most (Ringed seals are smaller, as are members of some lake-dwelling Common seal populations). Secondly, for their size they have among the biggest eyes of any pinniped, with the eyeballs being so big that they almost contact one another along the skull midline. Thirdly, the huge eyes appear to have forced other skull structures to become compressed or reduced: the frontal sinuses are strongly compressed and located further ventrally than is normal for seals; the nasal cavity has been forced into an unusually low position; and the brain has been displaced backwards. The jaw muscles, the ear region and the bones around the jaw joint are also modified relative to the condition in other seals, mostly in being smaller and weaker (Endo et al. 1998a, b, 1999). The foreflippers and their claws are larger and stronger in Baikal seals than is usual for seals, and the foreclaws are distinctive in being triangular in cross-section and in having a marked dorsal ridge at their distal end. It is inferred that these forelimb structures make Baikal seals better at making and maintaining breathing holes and grasping prey than other seals (Thomas et al. 1982).
 Baikal seals are thermophobic and pagophilic (that is, they avoid heat and like ice and snow), and their pups have white silky natal fur and are born on the ice that covers the lake between February and April. In these respects, Baikal seals are like the cold-adapted Ringed seal, Harp seal P. groenlandica and Ribbon seal P. fasciata, and as we’ll see this has implications for Baikal seal origins and history. Despite their small size, they are surprisingly long-lived, with males living to 52 and females to 56 (incidentally, Caspian seals are also long-lived, surviving to age 50). Average longevity for seals is about 20 years. Further remarkable is that female Baikal seals continue to reproduce while in their fourth decade.
Baikal seals are thermophobic and pagophilic (that is, they avoid heat and like ice and snow), and their pups have white silky natal fur and are born on the ice that covers the lake between February and April. In these respects, Baikal seals are like the cold-adapted Ringed seal, Harp seal P. groenlandica and Ribbon seal P. fasciata, and as we’ll see this has implications for Baikal seal origins and history. Despite their small size, they are surprisingly long-lived, with males living to 52 and females to 56 (incidentally, Caspian seals are also long-lived, surviving to age 50). Average longevity for seals is about 20 years. Further remarkable is that female Baikal seals continue to reproduce while in their fourth decade.
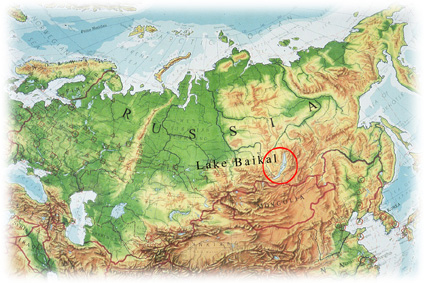
The great mystery, of course, concerns how the seals got into Lake Baikal: ultimately, we don’t yet know the proper answer, but that hasn’t stopped people from speculating. In fact it’s been said that more articles have been published on the biogeography of Lake Baikal’s seals than on the biogeography of all other pinnipeds combined. Even without the seals, Lake Baikal is a pretty remarkable lake, and would be better termed an inland sea. It’s 636 km long, nearly 80 km wide at its widest point, its average depth is 700 m and its maximum depth is 1.6 km. It is home to assorted endemic molluscs, crustaceans and nearly 60 fish species, and at least some of these animals are clearly of Arctic origin.

To date, two competing hypotheses have been proposed to explain the origins of the Baikal seals. Hypothesis 1 will be termed here the ‘Paratethyan hypothesis’, while hypothesis 2 will be termed the ‘Arctic origins hypothesis’.
The Paratethyan hypothesis
 The Paratethyan hypothesis accepts the conclusion, supported by some details of morphology, that Baikal seals, Ringed seals and Caspian seals all form a clade (the genus or subgenus Pusa*). Diverse phocine seals – some apparently resembling the extant Pusa** seals – are known to have inhabited Paratethys during the Miocene (Paratethys was a brackish inland sea that covered much of south-east Europe and south-west Asia during the Miocene) and, according to the Paratethyan hypothesis, it is phocines from this region that managed to invade the Caspian Sea, later getting as far east as Lake Baikal. During the Pliocene, Paratethys was linked to the Arctic Ocean via a seaway just west of the Urals, apparently, and accordingly it has been proposed that the Ringed seal of the Arctic Ocean descends from a phocine that migrated north from the Paratethys (Ray 1976, Grigorescu 1977) [adjacent image shows a Ringed seal].
The Paratethyan hypothesis accepts the conclusion, supported by some details of morphology, that Baikal seals, Ringed seals and Caspian seals all form a clade (the genus or subgenus Pusa*). Diverse phocine seals – some apparently resembling the extant Pusa** seals – are known to have inhabited Paratethys during the Miocene (Paratethys was a brackish inland sea that covered much of south-east Europe and south-west Asia during the Miocene) and, according to the Paratethyan hypothesis, it is phocines from this region that managed to invade the Caspian Sea, later getting as far east as Lake Baikal. During the Pliocene, Paratethys was linked to the Arctic Ocean via a seaway just west of the Urals, apparently, and accordingly it has been proposed that the Ringed seal of the Arctic Ocean descends from a phocine that migrated north from the Paratethys (Ray 1976, Grigorescu 1977) [adjacent image shows a Ringed seal].
* Whether the Pusa seals really share a single ancestor, however, has been contested and a recent DNA study found no support for the monophyly of Pusa: instead, the Baikal seal formed a clade, albeit weakly supported, with the Grey seal Halichoerus grypus (Palo & Väinölä 2006).
** Pusa pontica, from the Upper Miocene of Ukraine, has been referred to Pusa, but this is probably not correct.
Needless to say, there are problems with this hypothesis. Firstly, while it appears that the Caspian Sea is a relict of Paratethys (as is the Aral Sea), Lake Baikal appears way too far in the north-east for this to work, and where are all the connecting rivers and lakes that would be required in order to get seals from Paratethys all the way over to the Siberian interior? There are big lakes found between the Aral Sea and Lake Baikal, such as Lake Balkash, but they aren’t connected to the Paratethyan remnants, nor were they in the past. Secondly, the Paratethyan hypothesis requires that the phocines ancestral to Baikal seals and Ringed seals were animals of enclosed basins, relatively low latitudes and warm temperatures, and this is problematic given that Baikal and Ringed seals are thermophobic and pagophilic, with thickly-furred pups kept in snow dens. Furthermore, excepting the unusual inland populations, all phocines are oceanic, and not denizens of enclosed basins.
The Arctic origins hypothesis
The competing hypothesis posits that neither Baikal seals nor Ringed seals have descended from Paratethyan phocines that migrated south to north, but that all the pagopholic Pusa seals are of Arctic ancestry, and that seals got into Lake Baikal from the north. The fact that Baikal seals are thermophobic and pagopholic provides support for this model, as does the fact that many Baikal animals are apparently Arctic in origin.
Further support for the Arctic origins hypothesis comes from the fact that large ice-dammed lakes occupied central Siberia about 300,000 years ago. These apparently had connections with the Arctic Ocean and, via the Enisei-Angara river system, with Lake Baikal. Temporary downstream connections with the Caspian Sea also existed. This hypothesis better explains the ecology and life history of Baikal (and Caspian) seals, and is also superior to the Paratethyan hypothesis in that there are (and were) plausible dispersal routes that allowed the seals to get into the inland lakes. Worth noting is that Baikal seals occur in the rivers that flow north out of Lake Baikal even today. Particularly notable is a case where a seal was observed at the Irkutsk Dam on the Angara River, 400 km away from the lake (Thomas et al. 1982).
All in all, the Arctic origins hypothesis better fits the data and is the favoured hypothesis of recent authors (Deméré et al. 2003, Palo & Väinölä 2006). However, this isn’t the end of the debate. The degree of genetic divergence found amongst living phocines suggests – based on molecular clock inferences (which are always controversial and problematic) – that Baikal seals and other phocines diverged around 4 million years ago, during the late Pliocene. Why is this interesting? Because a divergence that occurred at this time doesn’t match with either the Paratethyan or the Arctic origin hypothesis.
 According to the former model, the ancestors of Baikal seals must have diverged from other phocines some time during the Miocene, and according to the latter model, Baikal seals evolved from a more northerly ancestor during the Pleistocene. In other words, if the genetic data is accurate, then the evolution of Baikal seals pre-dates the key event that is supposed to explain their distribution: the development of immense Siberian lakes connected both to the Arctic Ocean and to Lake Baikal. Palo & Väinölä (2006) therefore concluded that ‘the actual geographical conditions that would have facilitated the continental invasions in these times still remain undocumented and enigmatic’ (p. 70).
According to the former model, the ancestors of Baikal seals must have diverged from other phocines some time during the Miocene, and according to the latter model, Baikal seals evolved from a more northerly ancestor during the Pleistocene. In other words, if the genetic data is accurate, then the evolution of Baikal seals pre-dates the key event that is supposed to explain their distribution: the development of immense Siberian lakes connected both to the Arctic Ocean and to Lake Baikal. Palo & Väinölä (2006) therefore concluded that ‘the actual geographical conditions that would have facilitated the continental invasions in these times still remain undocumented and enigmatic’ (p. 70).
Huh. And there the story ends. For a previous seal-themed post see Swan-necked seals.
Coming soon: obscure dinosaurs of the Kimmeridge Clay. For the latest news on Tetrapod Zoology do go here.
Refs - -
Bonner, N. 1994. Seals and Sea Lions of the World. Blandford, London.
Deméré, T. A., Berta, A. & Adam, P. J. 2003. Pinnipedimorph evolutionary biogeography. Bulletin of the American Museum of Natural History 279, 32-76.
Endo, H., Sasaki, H., Hayashi, Y., Petrov, E. A., Amano, M. & Miyazaki, N. 1998a. Functional relationship between muscles of mastication and the skull with enlarged orbit in the Baikal seal (Phoca sibirica). Journal of Veterinary Medical Sciences 60, 699-704.
- ., Sasaki, H., Hayashi, Y., Petrov, E. A., Amano, M. & Miyazaki, N. 1998b. Macroscopic observations of the facial muscles in the Baikal seal (Phoca sibirica). Marine Mammal Science 14, 778-788.
- ., Sasaki, H., Hayashi, Y., Petrov, E. A., Amano, M., Suzuki, N. & Miyazaki, N. 1999. CT examination of the head of the Baikal seal (Phoca sibirica). Journal of Anatomy 194, 119-126.
Grigorescu, D. 1977. Paratethyan seals. Systematic Zoology 25, 407-419.
Kennedy, S., Kuiken, T., Jepson, P. D., Deaville, R., Forsyth, M., Barrett, T., van de Bildt, M. W. G., Osterhaus, A. D. M. E., Eybatov, T., Duck, C., Kydyrmanov, A., Mitrofanov, I. & Wilson, S. 2000. Mass die-off of Caspian seals caused by canine distempter virus. Emerging Infectious Diseases 6, 637-639.
King, J. E. 1983. Seals of the World. British Museum (Natural History), London.
Palo, J. U. & Väinölä, R. 2006. The enigma of the landlocked Baikal and Caspian seals addressed through phylogeny of phocine mitochondrial sequences. Biological Journal of the Linnean Society 88, 61-72.
Ray, C. E. 1976. Geography of phocid evolution. Systematic Zoology 25, 391-406
Smith, R. J., Hobson, K. A., Koopman, H. N. & Lavigen, D. M. 1996. Distinguishing between populations of fresh- and salt-water harbour seals (Phoca vitulina) using stable-isotope ratios and fatty acid profiles. Canadian Journal of Fisheries and Aquatic Science 53, 272-279.
Thomas, J., Pastukhov, V. & Petrov, E. 1982. Phoca sibirica. Mammalian Species 188, 1-6.
 Oh, and just for those who still don't accept the idea that a Golden eagle can kill a wolf...
Oh, and just for those who still don't accept the idea that a Golden eagle can kill a wolf...



































{kind=link}