Cadborosaurus and the Naden Harbour carcass: extant Mesozoic marine reptiles, or just bad bad science?
A recent meeting with Charles Paxton, a fisheries ecologist based at the Centre for Research into Ecological and Environmental Modelling at the University of St. Andrews, inspired me to think more about sea serpents [though note that I personally prefer the less loaded term ‘marine cryptid’*]. Charles is one of the few qualified scientists in the world publishing peer-reviewed papers on marine cryptids (Paxton 1998, 2004, Paxton & Holland 2005, Paxton et al. 2005), and while many technical zoologists have expressed an interest in this subject, Charles is pretty much the only one right now who is involved in detailed, empirical work on the subject. As usual, it’s funny how things pan out. For entirely unrelated reasons I’ve been writing about marine cryptids lately (more on that in future), plus I recently found myself writing about oarfishes and sea serpents in response to a post Reid Farmer put up on Steve Bodio’s Querencia blog (go here). During the week I was alerted to new Russian photos allegedly showing a marine cryptid carcass: alas, it is clear from the photos that the carcass is that of a Beluga Delphinapterus leucas.
* Cryptids are animals reported from anecdotal evidence and as yet unknown from museum-accessioned specimens.
While it might seem naïve or daft to think that aquatic cryptids, or sea serpents or whatever, really exist, there are actually reasonably good reasons to think that they do. Firstly, there is little doubt among scientists – I’d like to say ‘no doubt’ but that might be pushing it a bit too far – that large vertebrate species await discovery. By plotting species discovery rates over time to generate a species discovery curve, Paxton (1998) found that as many as 47 large open-water marine animals still await discovery, and new data caused him to up the figure to 51 a few years later (Paxton 2001). New study predicts a lower number (Paxton, pers. comm. 2006), though one that’s still surprisingly high. Raynal (2001) applied the same technique to cetacean discovery rates, and in this case the data suggested that 15 cetacean species await discovery. Solow & Smith (2005) also looked at this issue and concluded that about 10 large marine animal species await discovery, though they also found good statistical support for the possible presence of as many as 16 undiscovered species.
Secondly, such discoveries are not a hypothetical future event, but occur regularly, right here, right now. Megamouth shark Megachasma pelagios, discovered in the Pacific in 1976 and named in 1983*; Bandolero beaked whale (aka Peruvian or Lesser beaked whale) Mesoplodon peruvianus, known from specimens discovered 1975-1989, named as a new species in 1991; Spade-toothed whale M. traversii, named in 1874 but later sunk into synonymy: resurrected in 2002 when found to be synonymous with M. bahamondi (a supposedly new species named in 1995); Perrin’s beaked whale M. perrini known from specimens discovered 1975-1997 and named as a new species in 2002; Indonesian coelacanth Latimera menadoensis, discovered in 1997 and named in 1998; Omura’s whale Balaenoptera omurai, first collected in late 1970s and named in 2003; Australian snubfin dolphin Orcaella heinsohni, named in 2005.
* Megamouth sharks were discovered in the
Thirdly, the eyewitness evidence for marine cryptids is pretty good, in some cases being consistent across accounts and reported by trained observers who had a reputation to protect. They include military personnel and experienced naturalists. I could write thousands of words on such cases, so will hold off and only mention one: the
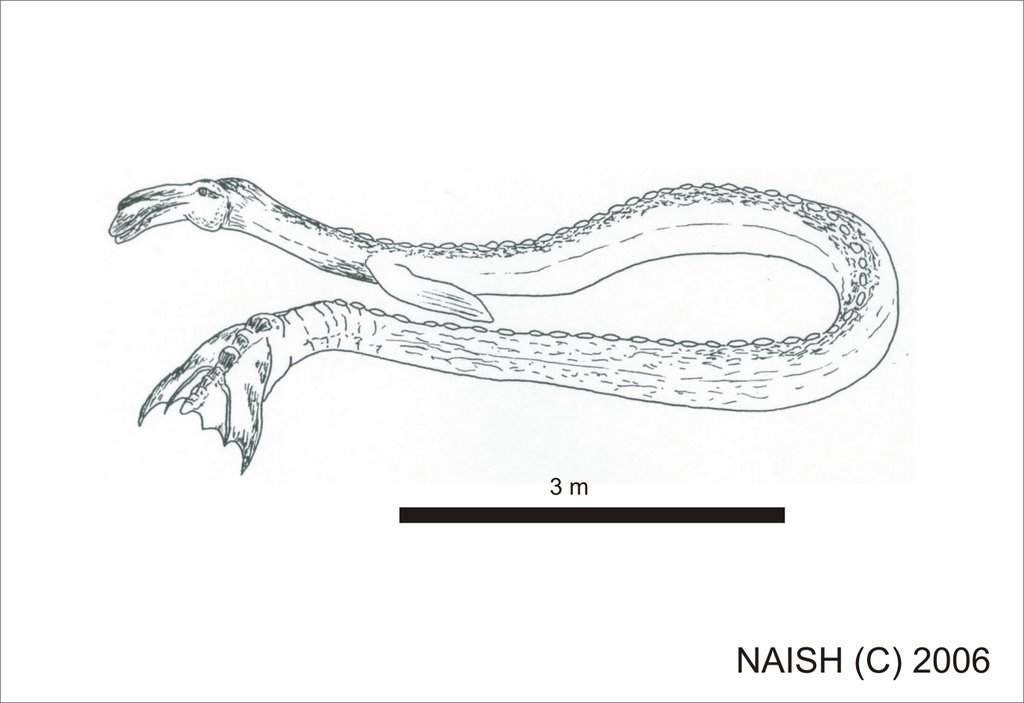
Of those marine cryptids that have attracted academic attention, perhaps the best known is that popularly dubbed ‘Caddy’ or ‘Cadborosaurus’, an enigmatic serpentine animal reported from the waters around the coasts of British Columbia. Named after Cadboro Bay, a sightings hotspot, Caddy sightings are impressively consistent, with most reports mentioning a large-eyed, horse-like or camel-like head (often with short horns), a long neck, serpentine body, a pair of flippers and a bifid, horizontal tail (LeBlond & Bousfield 1995). It is large, with estimated lengths ranging from 5 to 15 m. The animal’s body is often reported to sport brownish hair, and a serrated ridge is sometimes said to be present along the dorsal surface. Caddy has been described apparently pursuing and catching fish, and several accounts describe the animal snapping at, and even catching and eating, seabirds. It seems difficult to explain all of the sightings away as of misidentified cetaceans, elephant seals or large fish, and for now the identity of this creature remains a valid zoological mystery.

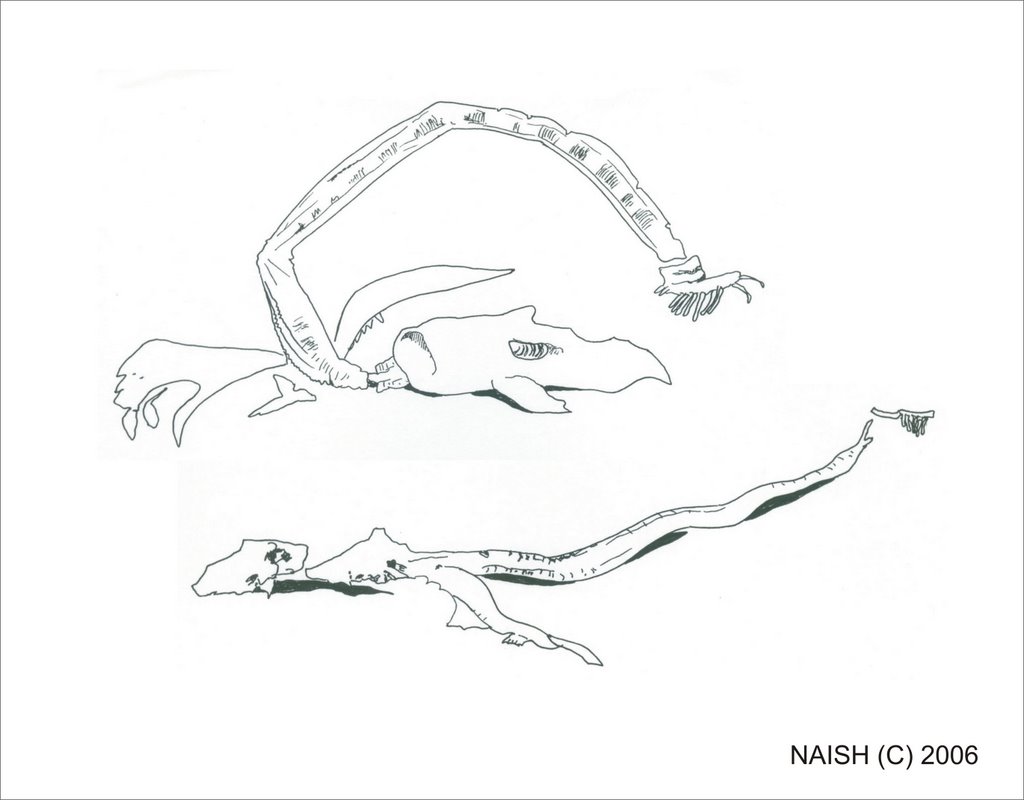
In 1992 Ed Bousfield, retired Research Associate at the Royal Ontario Museum, Toronto, and the Royal British Columbia Museum in Victoria, and Paul LeBlond, professor at the Department of Oceanography, University of British Columbia (Vancouver), announced some startling pieces of evidence - three photos, taken in 1937 on the flensing platform of a whaling station at Naden Harbour in the Queen Charlotte Islands, that depict a large serpentine carcass. Retrieved from the stomach of a sperm whale Physeter macrocephalus, the carcass is about 3 m long, long-bodied, and appears to have a camel-like head and a fluked tail. It obviously struck the whalers as unusual, otherwise they wouldn’t have gone to the trouble of setting it up on crates or photographing it. Suggestions that it is an elephant seal or baleen whale don’t seem realistic in view of certain of its features. These include an apparently symmetrical caudal fluke-like distal structure with a central series of knobs that appear to correspond to vertebrae (see close-up of photo and adjacent interpretative drawing below), and a camel-like ‘head’. The ‘body’ is elongate and serpentine and there appear to be pectoral flippers. While parts of the carcass were apparently retained and forwarded to the Pacific Biological Station at Nanaimo and/or the Royal British Columbia Museum, Victoria, no material remains today and the specimen is regarded as lost to science (Bousfield & LeBlond 1995, p. 9).

By combining observations on this carcass with eyewitness reports, Bousfield and LeBlond formally described Caddy as a new species, Cadborosaurus willsi and in 1995 devoted Supplement 1 of the new journal Amphipacifica to their paper on this taxon (Bousfield & LeBlond 1995). One of the
As mentioned above, while the number and quality of Caddy eyewitness accounts suggests that there may indeed be a new species awaiting discovery in the region, Bousfield and LeBlond’s proposal that the creature might be a living plesiosaur, and that such a species can be officially described and diagnosed based only on controversial photographic data, is highly problematical. Indeed an editorial that accompanied the publication of the Cadborosaurus description decried the lack of restraint employed by Bousfield and LeBlond, and strongly disagreed with the naming of the new species (Staude & Lambert 1995). These authors also noted that ‘Certainly it would have been preferable for [Bousfield and LeBlond] to publish in an independent journal, where neither served on the editorial board’ (p. 2). Aaron Bauer and Anthony Russell (1996) published a detailed critique of Bousfield and LeBlond’s paper, and I independently arrived at similar criticisms myself. The problems with Bousfield and LeBlond’s Cadborosaurus description can be grouped into three main areas.
1. How not to name a new species
Firstly, establishing a new species on the basis of a photo is just not acceptable: article 72(c)(v) of the International Code of Zoological Nomenclature states that the actual specimen figured or described, and not the illustration or description, must serve as the holotype. The name Cadborosaurus willsi, based on a photograph and not the specimen it depicts, therefore has no official standing and should be ignored. There are lots of other species names that have been coined in the cryptozoological literature in the absence of holotype specimens, and all are similarly nomina nuda that have no useful status. They include not only marine cryptids and relict hominids, but the Vietnamese snake Cryptophidion annamense, described from photos by Wallach and Jones (1992).

2. An erroneous identification
As discussed at depth in their book (LeBlond & Bousfield 1995), Bousfield and LeBlond favoured a reptilian identity for Cadborosaurus. Predominantly this is because they concluded that – in view of its elongate shape and occurrence in cold waters – Caddy must be poikilothermic. Jointed elements in the Naden Harbour specimen’s ‘tail pseudo-fluke’ (their term) were interpreted by Bousfield and LeBlond as being homologous with the hindlimb elements of Mesozoic marine reptiles, and they pointed in particular to a close similarity with the hindlimb of a pachypleurosaur (pachypleurosaurs are Triassic members of Sauropterygia, the reptile clade that includes plesiosaurs). Actually, while their figure (fig. 13C) definitely depicts the hindlimb of a pachypleurosaur, they identified it in their figure caption as the hindlimb of the plesiosaur Cryptoclidus. Furthermore, they mis-labelled most of the bones, identifying the tibia as the fibula, digit I as digit V and so on. [The image above shows a close-up of the carcass's 'head' together with an interpretative drawing, based on that provided by Bousfield & LeBlond (1995).]
The point of figuring a sauropterygian hindlimb was to show how the skeletal structure of the sauropterygian limb resembled the inferred bony structure of the
3. Inappropriate speculation, disturbing naivety
Overall, the Cadborosaurus description lacks the sort of academic rigour and restraint that is normal for taxonomic descriptions, so rather than bringing academic respectability to the study of Caddy it achieved just the opposite. With all due respect, Bousfield and LeBlond clearly did not do their homework when it came to some critical areas. If you are to seriously attempt to discuss the biology and morphology of any group of organisms in the academic literature, you must yourself consult the appropriate technical work that has previously been published: it is far from wise to rely on popular articles in magazines and encyclopaedic reference works written for laypeople. In their discussion of sauropterygians and basilosaurid cetaceans, for example, Bousfield and LeBlond cited popular and semi-popular works as their most authoritative references and apparently did not consult any of the technical literature on these subjects. Bauer & Russell (1996) noted that some of the terminology used by Bousfield and LeBlond implied ‘unfamiliarity with the literature’, and it’s telling that Bousfield and LeBlond referred to the archaic cetaceans Basilosaurus and Zeuglodon as if they were different, related genera, whereas in fact it has been widely acknowledged since the 1870s as least that Zeuglodon Owen is a junior synonym of the former.
 Also, one cannot seriously begin to write about plesiosaurs without consulting and citing articles produced by such workers as Welles, Riess, Massare, Cruickshank, Taylor, Brown, Bardet and Storrs: authors who have provided a wealth of functional, behavioural, taxonomical and morphological literature, much of which (e.g., hydrodynamics, ecology and feeding behaviours) would have been very pertinent to Bousfield and LeBlond’s speculations. Maybe if they had understood more of plesiosaur anatomy and functional morphology they would have sensibly reconsidered their proposal that Cadborosaurus was a possible plesiosaur descendant. It is actually very difficult to imagine a tetrapod MORE unlike a plesiosaur than is Cadborosaurus.
Also, one cannot seriously begin to write about plesiosaurs without consulting and citing articles produced by such workers as Welles, Riess, Massare, Cruickshank, Taylor, Brown, Bardet and Storrs: authors who have provided a wealth of functional, behavioural, taxonomical and morphological literature, much of which (e.g., hydrodynamics, ecology and feeding behaviours) would have been very pertinent to Bousfield and LeBlond’s speculations. Maybe if they had understood more of plesiosaur anatomy and functional morphology they would have sensibly reconsidered their proposal that Cadborosaurus was a possible plesiosaur descendant. It is actually very difficult to imagine a tetrapod MORE unlike a plesiosaur than is Cadborosaurus.
And having mentioned speculations, it has to be said that the number of speculations that Bousfield and LeBlond included within the paper are inappropriate for a work masquerading as a technical description, never mind the fact that those speculations were fantastic and logically flawed. It was proposed, for example, that the hair reported on Caddy might have a respiratory function (analogous to the hair-like growths seen on the frog Trichobatrachus), that the serpentine Cadborosaurus might somehow form a tuna-like body shape by bunching up the coils of its long body, that Caddy is viviparous and gives birth to large precocial babies, and that Caddy might be able to employ echolocation (Bousfield & LeBlond 1995). If these proposals sound nonsensical, or just extremely speculative and lacking in justification, that’s because they are.
So what now?
For me, the story of the
If the
 It’s also interesting that photos of an alleged ‘carcass’ very much like those taken at Naden Harbour were published as postcards in the 1930s (see adjacent interpretative drawings). Taken on the beach at
It’s also interesting that photos of an alleged ‘carcass’ very much like those taken at Naden Harbour were published as postcards in the 1930s (see adjacent interpretative drawings). Taken on the beach at
Finally, below is an excerpt from an article by Francois de Sarre and Michel Granger that mentions both Cadborosaurus and yours truly. The article was called Le serpent de mer existe bien! and appeared in Le Courrier of
----------------------------------------------------------------------------------------
Serpent de terre
En 1991, un cadborosaurus fut même signalé par des témoins dignes de foi près de Saanich Inlet comme ‘évoluant sur la terre ferme’.
Ce qui semblerait accréditer la filiation heuvelmansienne ‘mammalienne’ (issue du phoque ou du cétacé) plutôt que
L’absence systématique de queue appuie aussi cette descendance pinnipède, meme s’il est difficile d’imputer à une otarie la légende du serpent de mer.
Elle conviendrait beaucoup mieux, cepen-dant, pour décrire certains monstres de lacs tel Nessie, le célèbre monstre du
Et pourrait fournir, par un processus de déplacement ‘à nageoires sèches’, le passage dans le lack Okanagan, toujours en Colombie Brittanique, du monstre de Ogopogo.
Quand la nuit ou l’obscurité se fait à la tombée du jour, vous voyez une bête d’une vingtaine de mètres de long qui ondule à travers la campagne à la façon d’une chenille, raconte-t-on dans la région.
Un descendant des baleines?
Cette possibilité n’est pas exclue par le cryptozoologue brittanique Darren Naish quie met la description d’anneaux décollés de l’eau sur le compte de l’imagination des témoins. Mais, à notre avis, la question ne pourra être tranchée tant que d’autres éléments ne seront pas ajoutés au dossier du ‘monstre’. Par exemple, il n’est pas impossible qu’un filet de pêcheur résolve un jour l’énigme si ce n’est déjà fait. Les biologistes marins canadiens affi-chent un très bel optimsme dans ce sens. A quand un Cadborosaurus nageant dans un Marineland? Le fait d’envoyer un petit bathyscaphe dans
Les sceptiques n’ont qu’à bien se tenir. Pour notre part, nous pouvons affirmer: le légendaire serpent de mer existe bel et bein...
----------------------------------------------------------------------------------------
For previous blog posts on marine cryptids see Swan-necked seals and Gambo rides again. And for the latest news on Tetrapod Zoology please go here.
Refs - -
Bauer, A. M. & Russell, A. P. 1996. A living plesiosaur?: a critical assessment of the description of Cadborosaurus willsi. Cryptozoology 12, 1-18.
Bousfield, E. L. & LeBlond, P. H. 1992. Preliminary studies on the biology of a large marine cryptid in coastal waters of
- . & LeBlond, P. H. 1995. An account of Cadborosaurus willsi, new genus, new species, a large aquatic reptile from the Pacific coast of
Dash, M. 1993. The dragons of
LeBlond, P. H. & Bousfield, E. L. 1995. Cadborosaurus: Survivor from the Deep. Horsdal and Schubart Publishers (Victoria, B.C.).
Maede-Waldo, E. G. B. & Nicoll, M. J. 1906. Description of an unknown animal seen at sea off the coast of
Naish, D. 1997. Another Caddy carcass? The Cryptozoology Review 2 (1), 26-29.
Park, P. 1993. Beast from the deep puzzles zoologists. New Scientist 137 (1857), 16.
Paxton, C. G. M. 1998. A cumulative species description curve for large open water marine animals. Journal of the Marine Biologists Association,
- . 2001. Predicting pelagic peculiarities: some thoughts on future discoveries in the open seas. In Heinselman, C. (ed) Dracontology Special Number 1: Being an Examination of Unknown Aquatic Animals. Craig Heinselman (
- . 2004. Giant squids are red herrings: why Architeuthis is an unlikely source of sea monster sightings. The Cryptozoology Review 4 (2), 10-16.
- . &
- ., Knatterud, E. & Hedley, S. L. 2004. Cetaceans, sex and sea serpents: an analysis of the Egede accounts of a “most dreadful monster” seen off the coast of
Raynal, M. 2001. Cryptocetology and mathematics: how many cetaceans remain to be discovered? In Heinselman, C. (ed) Dracontology Special Number 1: Being an Examination of Unknown Aquatic Animals. Craig Heinselman (
Solow, A. R. & Smith, W. K. 2005. On estimating the number of species from the discovery record. Proceedings of the Royal Society B 272, 285-287.
Staude, C. P. & Lambert, P. 1995. Editorial… an opposing view. Amphipacifica 1 (Supp. 1), 2.
Wallach, V. and Jones, G. S. 1992. Cryptophidion annamense, a new genus and species of cryptozoic snake from
White, W. T., Muhammad Adrim, F. & Sumadhiharga, K. 2004. A juvenile megamouth shark Megachasma pelagios (Lamniformes: Megachasmidae) from northern
Labels: cryptozoology, marine reptiles, sea serpents
posted by Darren Naish at 9:30 PM
![]()
![]()


12 Comments:
I can give another reason to at least be suspicious of this "Caddy carcass." You said that the whalers claim the specimen was "retrieved from the stomach of a sperm whale Physeter macrocephalus," and it was "about 3 m long." As far as I can tell from my library and some web searches, sperm whales don't eat large vertebrate prey at all. Small vertebrates, yes -- one source even said they sometimes eat seals. But a fish (or anything else) 3m long? And swallowed whole, or at least intact enough to recognize and reassemble? Sounds a bit odd to me.
I'd also suggest a review of Dr. Bernard Heuvelmans' superb book In the Wake of the Sea-Serpent (1968). He spends an entire chapter on cryptic carcasses that were washed up on various beaches, and succeeds in showing that several of these "sea serpents" were probably badly decayed basking sharks. Glen Kubans' look at the Zuiyo Maru "sea serpent" is also a good and relevant read.
To me it seems the picture shows a half digested tentacle of the Collosal Squid (or perhaps the Giant Squid), with the presumed "head" being the tentacle club and the "vertebrae" being the suckers. It is what one would expect to find in the stomach of a sperm whale, but perhaps its rather large size made it an object interesting enough to photograph.
Great entry! And I can really sympathise with your post-doc blues. I did my PhD on 1st Millennium graves on an island in the Baltic...
The Blogger upgrade thing means I can't use my account to comment here, but you'll find me at saltosobrius.blogspot.com.
Martin
Darren, for some reason every time I ask for the comment window my browser crashes. I don't know if other people are having this problem (I'm using firefox).
Darren, I must say that you have the most disturbingly logical approach to Cryptozoology that I've seen on the internet. I've always been pretty baffled by LeBlond and Bousfield's conclusions, wouldn't a reptilian poikilotherm in 40-50 degree water be pretty much unfunctional? And why would a plesiosaur or any other reptile suddenly decide to undulate vertically anyways? Your post confirmed my suspicions and then some (I haven't read the paper), I had no idea that the "science" was that shoddy.
I really enjoyed your point about the Fircom carcass, I'd always wondered if the "carcass" was just some piece of rubbish. One thing that always confused me was how the "head" looks so different in the two pictures, unfeatured in the angled pic (on the site), and looking more biological in the lengthwise pic. Unfortunately that other picture is pretty hard to find on the internet. I think that would imply that the picture is unclear enough that pareidolia is taking place and details are being added to where they aren't.
In addition to Paxton, Bruce Champagne may still be working on his sea serpent, er, marine cryptids, but I'm not sure what shape it's in or what is different from the "Dracontology" publication a few years back.
-Cameron
On sperm whale diets, I take your point that swallowing a 3-m-long vertebrate might seem unlikely. However, I consulted the technical literature and there are numerous reports of BIG intact animals (vertebrates and invertebrates) being recovered from sperm whale stomachs. Of 32 squid recovered from one whale, the average size was 2.2 m (Clarke 1954, p. 313) and architeuthids (giant squid) recovered from sperm whale stomachs have been stated to have an average mantle length (viz, not total length) of over 1 m (Clarke et al. 1993). An intact squid 10.49 m long and 184 kg in weight was recovered from a 14.6 m long whale killed in the Azores (Clarke 1955, p. 355). What about vertebrates? Sperm whales are recorded as having eaten sharks 2.5 and 3 m long, including basking and blue sharks (Norman & Fraser 1938, Caldwell et al. 1966). And yes, there are those stories about sperm whales swallowing people, but I don't know what to make of those.
In conclusion, sperm whales certainly are capable of swallowing surprisingly large animals, and they do so, and they swallow their prey whole.
On marine cryptids, I know Heuvelmans 1968 well, and of course have been consulting it lately. But it wasn't really relevant to the Caddy post, so I didn't need to cite it. Many of the carcasses he reviews have since been re-evaluated, for which you need to see the series of articles on mysterious marine carcasses produced by Roesch (1997, 1998a, b, 1999). I'm also familiar with the Zuiyo Maru case and in fact helped Glen in his research (check the acknowledgements).
Thanks for your thoughtful comments.
To those interested, I removed the mention of Conan-Doyle given that it caused such a lot of extraneous discussion over at Pharyngula.
Refs - -
Caldwell, D. K., Caldwell, M. C. & Rice, D. W. 1966. Behavior of the sperm whale, Physeter catodon L. In Norris, K. S. (ed) Whales, Dolphins and Porpoises. University of California Press (Berkeley & Los Angeles), pp. 678-717
Clarke, M. R., Martins, H. R. & Pascoe, P. 1993. The diet of sperm whales (Physeter macrocepahlus Linnaeus 1758) off the Azores. Philosophical Transactions of the Royal Society of London B 339, 67-82.
Clarke, R. 1954. Whales and seals as resources of the sea. Norsk Hvalgangst-tid. 43, 301-317.
- . 1955. A giant squid swallowed by a sperm whale. Norsk Hvalgangst-tid. 44, 353-357.
Norman, J. R. & Fraser, F. C. 1938. Giant Fishes, Whales and Dolphins. University of California Press, Berkeley.
Roesch, B. S. 1997. A review of alleged sea serpent carcasses worldwide (part one – 1648-1880). The Cryptozoology Review 2 (2), 6-27.
- . 1998a. A review of alleged sea serpent carcasses worldwide (part two – 1881-1896). The Cryptozoology Review 2 (3), 25-35.
- . 1998b. A review of alleged sea serpent carcasses worldwide (part three – 1897-1906). The Cryptozoology Review 3 (1), 27-31.
- . 1999. A review of alleged sea serpent carcasses worldwide (part four – 1907-1924). The Cryptozoology Review 3 (3), 15-22.
Where does one report an actual Caddy sighting? And is anyone interested I wonder? I had a close encounter with one several years ago when camping in a remote area of west coast Vancouver Island.
Hi Jim
The best people to report a sighting to are probably those at the British Columbia Scientific Cryptozoology Club. A contact email can be found at their website... http://www.bcscc.ca/index.htm. Certainly there are interested people.
The head reminds me of a dugong. Maybe it was just a dugong status that's just blown off course some how.
(A pretty big "Somehow" but htere you go)
I think the reason nobody has found a corpse is because of the structure of the body ? It doesn't seem like something that would wash up like most things in BC " fish, whales, sharks" it's like a snake and after all the years on the beach here in BC I've seen nothing similar washed up. I think they may be bottom feeders and surface only in distress or mating which would maybe explain why the are so hard to spot, many sightings involve more then one so perhaps they are mating. Fish I have seen washed up would support the idea of a large bottom feeder with teeth much different then whales and sharks. Halibut I would guess is it's main diet. I'm no scientist just a beach walker.
There is more pictures of this creature published in a book printed in Canada sometime in the 50's/60's, I have been looking for it scince I was a kid and signed it out of the Port Mc Neil school libary when I lived there in the eighties. The pictures are on docks and clearer unlike these they are much better and of a undecomposed creature. surprised no one has put them up yet. In my opinion from walking on the beach for thirty years this creature is a bottom feeder based on the fish I've seen half consumed washed up on the beach with much different bites then sharks and whales. Perhaps we never find corpses because of the nature of bottom feeders here in BC, they consume everything organic very quickly and maybe when we see them on the surface it is because they are mating or in distress. Halibut I would say is their main diet. Just a guess
Great article! Very Professional.
I hope some definitive evidence for Caddy shows up. The whole thing is fascinating.
Post a Comment
<< Home