Frame 352, and all that
 For some time now I’ve been toying with the idea of writing a blog post about sasquatch,
For some time now I’ve been toying with the idea of writing a blog post about sasquatch, In the interests of hypothesis testing, I finally decided: what the hell. My hypothesis is: will writing about sasquatch negatively affect my career prospects? Well, let’s test the hypothesis. Let me state from the start that I do not “believe” in sasquatch, nor am I planning to promote either an anti-sasquatch, or a pro-sasquatch, point of view. What I have learnt from research on this area is that – contrary to the assertions of some – the evidence for sasquatch is, at the very least, scientifically interesting and worthy of investigation.
While purported evidence for the supposed reality of sasquatch continues to attract strong criticism, more interesting in my view is that a number of academically qualified primatologists have recently gone on record in stating that the evidence for sasquatch is scientifically compelling. These people do not only include well-known sasquatch proponents, such as the late Grover Krantz (1931-2002) of
Here I am going to discuss one particular piece of evidence for sasquatch: the Patterson-Gimlin film. This is that famous short piece of film that you’ve probably seen on TV many times: it depicts what appears to be an obviously female sasquatch striding across a clearing from left to right [for M. K. Davis’ stabilised version of the film go here]. You’ve probably heard that the film has been revealed to be a hoax. Well, sorry, that ain’t true.
 On
On
 1. TAS looks genuine. Its coat is glossy, conforms to the underlying contours, muscular bulges, joints and other structures in the body, and looks realistic compared to living mammals. What appears to be a shallow parting extends axially along the spine and between the buttocks [in adjacent image, note the demarcated buttocks and apparent wear on the buttocks]. As TAS moves, its muscles (in its legs and elsewhere) can be seen to bulge and flex beneath the fur as they do in living mammals. TAS’s gait is fluid and natural and it differs in subtle details of posture and proportion from humans (see points 2 and 3). Its toes are seen to lift at one point. Its large breasts bounce and sway in a manner which looks realistic compared to how unsupported human breasts move during locomotion. It is also intriguing that TAS’s compliant gait and protruding heel match features reported by eyewitnesses (see point 2). High quality enlargements have been published of key frames from the footage several times (e.g. Bayanov 1997, Murphy et al. 2004) so it is easy to check all of these assertions. Put in its simplest terms: despite claims to the contrary, TAS looks realistic.
1. TAS looks genuine. Its coat is glossy, conforms to the underlying contours, muscular bulges, joints and other structures in the body, and looks realistic compared to living mammals. What appears to be a shallow parting extends axially along the spine and between the buttocks [in adjacent image, note the demarcated buttocks and apparent wear on the buttocks]. As TAS moves, its muscles (in its legs and elsewhere) can be seen to bulge and flex beneath the fur as they do in living mammals. TAS’s gait is fluid and natural and it differs in subtle details of posture and proportion from humans (see points 2 and 3). Its toes are seen to lift at one point. Its large breasts bounce and sway in a manner which looks realistic compared to how unsupported human breasts move during locomotion. It is also intriguing that TAS’s compliant gait and protruding heel match features reported by eyewitnesses (see point 2). High quality enlargements have been published of key frames from the footage several times (e.g. Bayanov 1997, Murphy et al. 2004) so it is easy to check all of these assertions. Put in its simplest terms: despite claims to the contrary, TAS looks realistic. 2. TAS walks with a compliant gait, and not with the same striding knee-locking gait of humans [adjacent image shows frame 352, the most famous and oft-shown part of the film]. Its knee is never fully straightened in its step cycle, even in the supporting phase. Its arms swing slightly more than those of humans, and its hands and wrists are held supinated and slightly flexed with the fingers curved (this is unlike the normal hand posture of humans). It’s clearly possible that all of these features could be faked by a knowledgeable human, and Daegling & Schmitt (1999) argued that the gait and speed used by TAS can be reproduced by humans. That person would, however, have to not only conform physically to the dimensions of TAS (see point 3), but would also have to be very good at walking with an unusual gait which is practised so well that it has convinced experts in biomechanics and primate anatomy (see point 4). That person would also have to be an expert, or at least supervised by one, on the eyewitness data (which describes identical points of posture and morphology). It is unlikely that such a person exists and/or was available to Patterson and/or Gimlin in 1967, and extensive biographical research on Patterson and Gimlin and their friends and colleagues has failed to uncover the existence of any such person.
2. TAS walks with a compliant gait, and not with the same striding knee-locking gait of humans [adjacent image shows frame 352, the most famous and oft-shown part of the film]. Its knee is never fully straightened in its step cycle, even in the supporting phase. Its arms swing slightly more than those of humans, and its hands and wrists are held supinated and slightly flexed with the fingers curved (this is unlike the normal hand posture of humans). It’s clearly possible that all of these features could be faked by a knowledgeable human, and Daegling & Schmitt (1999) argued that the gait and speed used by TAS can be reproduced by humans. That person would, however, have to not only conform physically to the dimensions of TAS (see point 3), but would also have to be very good at walking with an unusual gait which is practised so well that it has convinced experts in biomechanics and primate anatomy (see point 4). That person would also have to be an expert, or at least supervised by one, on the eyewitness data (which describes identical points of posture and morphology). It is unlikely that such a person exists and/or was available to Patterson and/or Gimlin in 1967, and extensive biographical research on Patterson and Gimlin and their friends and colleagues has failed to uncover the existence of any such person.3. TAS is physically large and with proportions that appear to be unlike those of our species. Its intermembral index (the ratio of humerus + radius length to femur + tibia length) is between 80 and 90, whereas in our species it averages 72*, and its breadth across its shoulders is about 35% of its total height. Krantz (1999) asserted that some humans (including inuit people) have a shoulder breadth that exceeds 30% of total height (this is apparently not the case in people that exceed 2 m in height), and that other data also indicates that the creature exceeds in torso width any human. Krantz (1999) concluded on the basis of this evidence ‘I can confidently state that no man of that stature is built that broadly’. However, Daegling & Schmitt (1999) challenged this torso-breadth data, and argued that the estimates do overlap with that from tall humans.
* In chimps and gorillas the intermembral indices are 106 and 117, respectively. TAS therefore seems intermediate between chimps and humans with regard to this feature.
Patterson and Gimlin photographed, and took casts from, a trackway which (they asserted) was made by TAS. These photographs and tracks survive today and both (i) appear genuine* and (ii) correspond with the details of TAS’s size and gait as seen in the footage. Average track length was 36.8 cm, and because the full length of TAS’s foot sole can be seen in several frames, the sole : total height ratio of about 1 : 5 gives a rough height of 184 cm. A similar height has been estimated by triangulation, by working out how the stride length used by TAS matches with humans of various statures, and by other methods.
* That is, like other ‘good’ sasquatch prints, they appear to have been made by a large, very heavy hominid with a flexible foot that exhibits several consistent anatomical novelties.
 4. Several workers experienced with primate biomechanics and locomotion have examined the footage, and in several cases have published comments on it. All have concluded either that the film is genuine and depicts a non-human primate, or have admitted that their examination was inconclusive.
4. Several workers experienced with primate biomechanics and locomotion have examined the footage, and in several cases have published comments on it. All have concluded either that the film is genuine and depicts a non-human primate, or have admitted that their examination was inconclusive.Dmitri Donskoy [Chief of the Chair of Biomechanics at the USSR Central Institute of Physical Culture,
D. W. Grieve [Reader in Biomechanics, Royal Free Hospital School of Medicine, London] concluded ‘The possibility of fakery is ruled out if the speed of the film was 16 or 18 fps [as mentioned above, it was apparently filmed at 16 fps]. In these conditions a normal human being could not duplicate the observed pattern, which would suggest that the sasquatch must possess a very different locomotor system to that of man’.
Grover Krantz, well known as an advocate of sasquatch but nonetheless still an experienced and qualified anthropologist, argued that the creature’s size, proportions and gait demonstrated its genuine nature, concluding ‘there is no possibility that the film can be a man in a fur suit’. Bayanov (1997) cited views from several Russian biomechanists who thought that the creature’s gait could not be reproduced by a human. Jürgen Konczak [associate professor in the
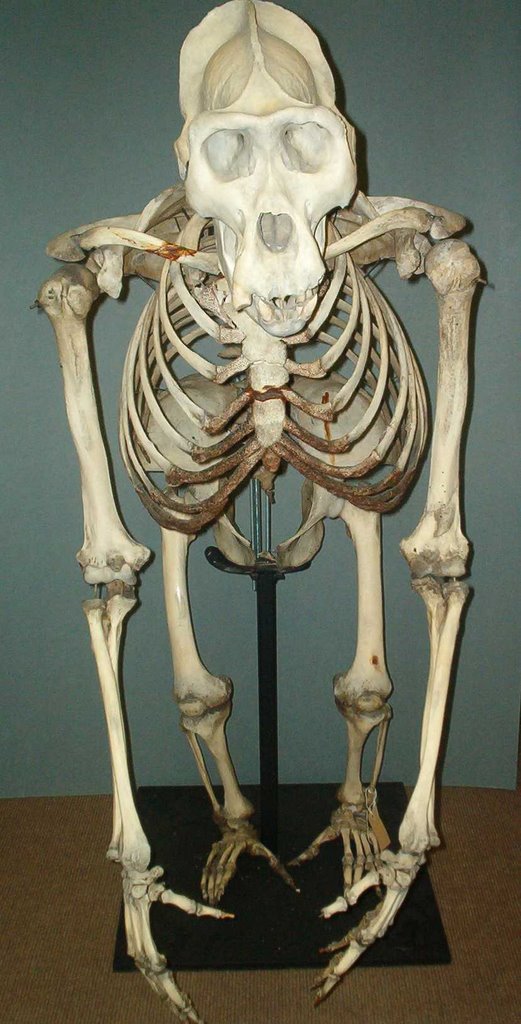 In view of this large number of ‘positive’ interpretations, most of which come from authoritative, technically qualified experts who do not have any axe to grind on the issue of sasquatch, what evidence has been marshalled by those who assert that the film is faked? To date, none. No analysis has been performed which shows that the creature can be explained as a man in a suit. Published objections have either asserted that the animal walks in a manner ‘consistent in general terms with the bipedal striding gait of modern man’ (Napier 1974), or have pointed to the presence of furry breasts, the presence of a sagittal cranial crest in a female, or the presence of breasts in a creature without female-like hips and a waist, as problems showing that the film must have been faked. These objections are all clearly erroneous (e.g. it is difficult to be confident that furry breasts are somehow impossible – while many primates do sport naked pectoral skin around their nipples and areolae, human breasts are hairy, it’s just that the hairs are very small and thin; sagittal crests are size-related, and only absent in the females of most hominid species because females do not match adult males in the size of their cranial musculature; broad hips and a waist are characters of our species, and not of other hominids or primates [gorilla skeleton at left]). Napier’s objections were vague and have not been supported by other workers experienced in biomechanics.
In view of this large number of ‘positive’ interpretations, most of which come from authoritative, technically qualified experts who do not have any axe to grind on the issue of sasquatch, what evidence has been marshalled by those who assert that the film is faked? To date, none. No analysis has been performed which shows that the creature can be explained as a man in a suit. Published objections have either asserted that the animal walks in a manner ‘consistent in general terms with the bipedal striding gait of modern man’ (Napier 1974), or have pointed to the presence of furry breasts, the presence of a sagittal cranial crest in a female, or the presence of breasts in a creature without female-like hips and a waist, as problems showing that the film must have been faked. These objections are all clearly erroneous (e.g. it is difficult to be confident that furry breasts are somehow impossible – while many primates do sport naked pectoral skin around their nipples and areolae, human breasts are hairy, it’s just that the hairs are very small and thin; sagittal crests are size-related, and only absent in the females of most hominid species because females do not match adult males in the size of their cranial musculature; broad hips and a waist are characters of our species, and not of other hominids or primates [gorilla skeleton at left]). Napier’s objections were vague and have not been supported by other workers experienced in biomechanics.David Daegling [associate professor of anthropology at Yale University] and Daniel Schmitt [assistant professor in the Department of Anthropology, Duke University Medical Center] published an article in Skeptical Inquirer in which they argued that TAS’s size and style of gait can be reproduced by people. They were still unable to assert that it was fake however, concluding ‘Based on our analysis of gait and problems inherent in estimating subject dimensions, it is our opinion that it is not possible to evaluate the identity of the film subject with any confidence’.
Multiple claims have been made that the footage was faked by a known individual, and that this individual has provided a death-bed confession, or something like that. It has been easy to knock down all of these claims and show them to be fabrications (e.g. Coleman 2003, Murphy et al. 2004, Vella 2004, Perez 2005).
 5. In view of these observations, it is difficult to take seriously claims that TAS is actually some tall guy in a gorilla costume. Even today there is no maker of fake/synthetic fur, or of animal costumes, who can reproduce something this realistic, nor are there any suits which look so realistic, which allow the mimicry of moving musculature and breasts, and which are anatomically accurate compared to living primates. Two serious attempts have been made to reproduce the film using a man in a specially designed suit: one for the BBC TV series ‘The X Creatures’ [image at left]; the second for Kal Korff’s documentary ‘The Making of Bigfoot’. In both instances the resulting attempt to discredit the Patterson-Gimlin film backfired: their results look like a man in a monkey suit, and in no way come even close to resembling TAS in the 1967 Patterson-Gimlin film. Several special effects experts have been consulted on how possible it might be to reproduce what’s seen in the footage (this is particularly relevant as there have been repeated claims that someone in the Hollywood special effects community manufactured a suit for Patterson), including John Chambers [designer of the ape costumes seen in ‘Planet of the Apes’]. While some have claimed that the construction of a suit matching what’s seen in the Patterson film would be easy or possible, I am troubled by the fact that no-one has yet replicated it. At least some special effects people have stated that the creature seen in the footage exceeds in accuracy and realism the special effects available to workshops today, let alone those existing in 1967.
5. In view of these observations, it is difficult to take seriously claims that TAS is actually some tall guy in a gorilla costume. Even today there is no maker of fake/synthetic fur, or of animal costumes, who can reproduce something this realistic, nor are there any suits which look so realistic, which allow the mimicry of moving musculature and breasts, and which are anatomically accurate compared to living primates. Two serious attempts have been made to reproduce the film using a man in a specially designed suit: one for the BBC TV series ‘The X Creatures’ [image at left]; the second for Kal Korff’s documentary ‘The Making of Bigfoot’. In both instances the resulting attempt to discredit the Patterson-Gimlin film backfired: their results look like a man in a monkey suit, and in no way come even close to resembling TAS in the 1967 Patterson-Gimlin film. Several special effects experts have been consulted on how possible it might be to reproduce what’s seen in the footage (this is particularly relevant as there have been repeated claims that someone in the Hollywood special effects community manufactured a suit for Patterson), including John Chambers [designer of the ape costumes seen in ‘Planet of the Apes’]. While some have claimed that the construction of a suit matching what’s seen in the Patterson film would be easy or possible, I am troubled by the fact that no-one has yet replicated it. At least some special effects people have stated that the creature seen in the footage exceeds in accuracy and realism the special effects available to workshops today, let alone those existing in 1967. There’s a lot more that could be said on this subject, but I’ll leave it at that. I have not discussed Roger Patterson’s personal circumstances (relevant to claims that he faked the footage for money or fame), nor have I touched on the interesting story of what a farce Patterson and Gimlin’s eventual development and treatment of the film was. As Richard Greenwell (1942-2005) – former secretary of the International Society of Cryptozoology – said to me in a letter of March 2000: ‘In the big picture it matters little if Bigfoot exists or not; what matters is that proper procedure be followed in examining such evidence – or any evidence’.
UPDATE (29-11-2006): Loren Coleman has written a blog post about this one - see Napier, Naish, and Frame 352. For the latest news on Tetrapod Zoology please go here.Refs - -
Bayanov, D. 1997.
Daegling, D. J. & Schmitt, D. O. 1999. Bigfoot’s screen test. Skeptical Inquirer May/June 1999, 20-25.
Coleman, L. 2003. Bigfoot! The True Story of Apes in
Krantz, G. S. 1999. Bigfoot Sasquatch Evidence. Hancock House,
Murphy, C. L., Green, J. & Steenburg, T. 2004. Meet the Sasquatch. Hancock House,
Napier, J. 1974. Bigfoot. Readers
Perez, D. 2004. In defence of the Patterson-Gimlin film. Fortean Times 192, 36-37.
Vella, P. 2004. J’accuse. Animals & Men 34, 42-48.
Labels: cryptozoology, mammalogy, mammals
posted by Darren Naish at 12:46 AM
41 comments
![]()
![]()






















{kind=link}