Giant hoatzins of doom
 Inspired by the recent description of a new and exciting phorusrhacid – a giant predatory South American landbird – I have lately been producing various blog posts on members of this group, as you’ll know if you’re a regular reader. See Terror birds and More on phorusrhacids. There is lots more to say: I am planning also to blog about phorusrhacid skull and hand anatomy, and about their alleged survival into near-modern times. The horrible danger is that the distractions that constantly arise will cause me to veer off at a tangent, and already the paper on the British dinosaurs of the Kimmeridge Clay that Dave Martill, Sarah Fielding and I have finally had published has me wanting to move on to something else. I am also desperate to blog about the lions of Chobe National Park (Botswana), as their ability to kill adult elephants has finally been filmed and is due to be screened on TV next week. Furthermore, there is that AMAZING discovery of a bottlenose dolphin with a perfect pair of miniature hind-flippers (go here), and there is that Mantellisaurus thing. Stay tuned.
Inspired by the recent description of a new and exciting phorusrhacid – a giant predatory South American landbird – I have lately been producing various blog posts on members of this group, as you’ll know if you’re a regular reader. See Terror birds and More on phorusrhacids. There is lots more to say: I am planning also to blog about phorusrhacid skull and hand anatomy, and about their alleged survival into near-modern times. The horrible danger is that the distractions that constantly arise will cause me to veer off at a tangent, and already the paper on the British dinosaurs of the Kimmeridge Clay that Dave Martill, Sarah Fielding and I have finally had published has me wanting to move on to something else. I am also desperate to blog about the lions of Chobe National Park (Botswana), as their ability to kill adult elephants has finally been filmed and is due to be screened on TV next week. Furthermore, there is that AMAZING discovery of a bottlenose dolphin with a perfect pair of miniature hind-flippers (go here), and there is that Mantellisaurus thing. Stay tuned. While I’m here I want to note some recent updates I’ve made to some articles. Firstly, thanks to comments that have been added by anonymous mammalogists (PLEASE leave your name when you leave a comment), I’ve done some minor updating to The first new European mammal in 100 years? We are now at 31 ‘100 year’ mammals. Secondly, inspired by some questions from Mark Abuys, I have also added some new comments to Graeme’s Pleistocene megafrog. Mark was interested in my mention of the carn-pnay, a crypto-frog from
To return to phorusrhacids...... in the previous posts I at least alluded to ideas about their affinities: it is universally agreed that their closest living relatives are the South American seriemas (or cariamids), and it is furthermore agreed that two fossil groups, the bathornithids of Eocene-Miocene North America and the idiornithids of Eocene-Oligocene Europe, are also close relatives of both seriemas and phorusrhacids. Several features unite these birds (Mayr 2002). They all have a strongly hooked bill, a simple, block-like hypotarsus (a site of ligament attachment on the posterior surface of the tarsometatarsus), distinctively proportioned toe bones, and a laterally compressed, strongly curved and sharp-tipped claw on the second toe (discussed previously here). All of these birds – idiornithids, bathornithids, cariamids and phorusrhacids – form a clade termed the Cariamae Fürbringer, 1888, and conventionally they’ve been regarded as part of Gruiformes, the group that includes rails, cranes, trumpeters and several other groups. A few other fossil groups have been suggested to be part of Cariamae, like the cunampaiids of Eocene Argentina.
Are cariamaens really close allies of rails and cranes? Storrs Olson (1985, p. 143) wrote that the classification of cariamaens within Gruiformes was ‘largely by default, as they do not clearly seem to belong in any other order’, and there is a long-running disagreement among ornithologists as to whether gruiforms are a natural group or not. We’ll look at this issue in the next post.
The South American landbird hypothesis
If you’re a regular reader of this blog you’ll know that I’m a big fan of ‘alternative’ theories on phylogenetic relationships (see, for example, We flightless primates). While I always find these ideas interesting, note the caveat that I do not necessarily endorse them: in fact they often turn out to be poorly supported or spurious. As it happens, one of my favourite ‘alternative’ theories involves cariamaens: it is the South American landbird hypothesis.
 Based predominantly on the morphology of the carpometacarpus, some ornithologists have proposed that cariamaens are closely related to the Hoatzin* Opisthocomus hoazin**, that bizarre folivorous, arboreal bird that (uniquely among birds) practices foregut fermentation. In contrast to those of most other neornithines, the carpometacarpi of cariamaens and hoatzins possess a particularly broad, strongly bowed third metacarpal. This is also true of turacos and some cuckoos (indeed, many ornithologists have proposed that hoatzins might be close allies of turacos and/or cuckoos).
Based predominantly on the morphology of the carpometacarpus, some ornithologists have proposed that cariamaens are closely related to the Hoatzin* Opisthocomus hoazin**, that bizarre folivorous, arboreal bird that (uniquely among birds) practices foregut fermentation. In contrast to those of most other neornithines, the carpometacarpi of cariamaens and hoatzins possess a particularly broad, strongly bowed third metacarpal. This is also true of turacos and some cuckoos (indeed, many ornithologists have proposed that hoatzins might be close allies of turacos and/or cuckoos).
* Like ‘fossa’ and ‘sifaka’, ‘hoatzin’ is one of those words that is, apparently, not pronounced the way in which it is written. Some sources state that it is properly pronounced ‘watson’.
** That’s not a typo. Furthermore, the name Opisthocomus cristatus, used by some authors relatively recently (e.g., Chatterjee in The Rise of Birds) was coined (so far as I can tell) by Johann Illiger in 1811 and thus post-dates Statius Muller’s 1776 publication of the name Phasianus hoazin (that’s right - the hoatzin was originally described as a type of pheasant).
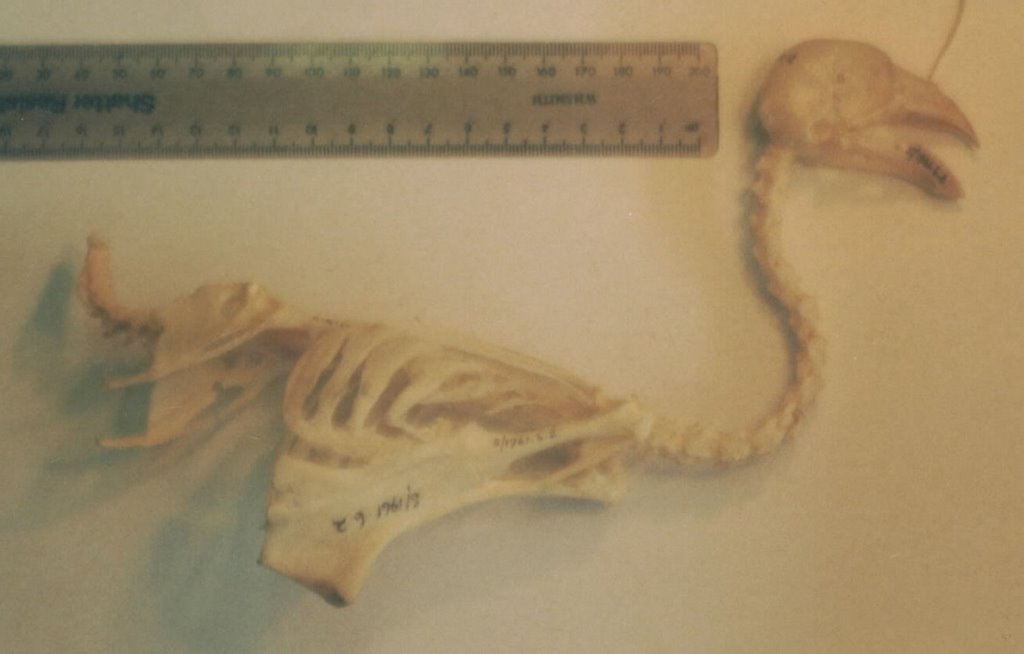
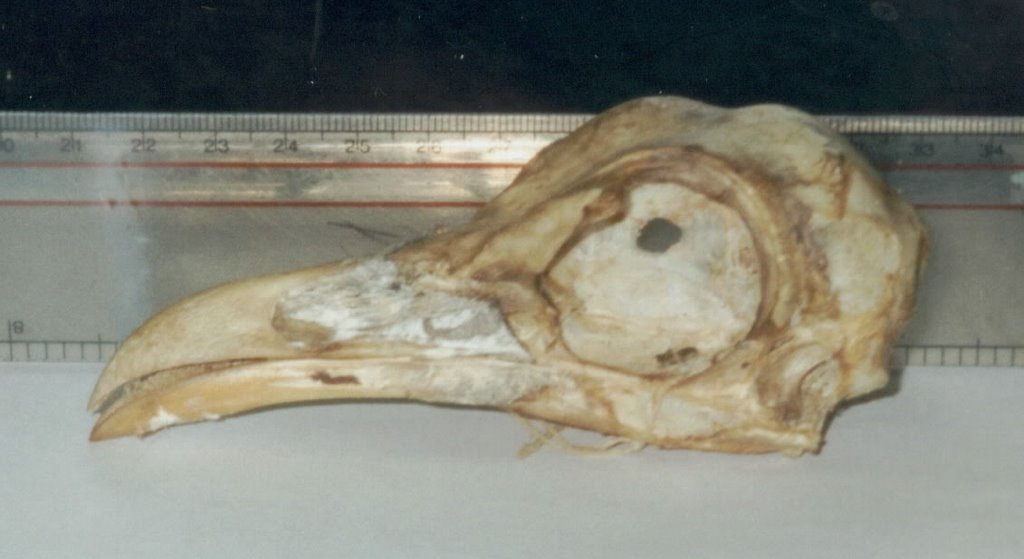 Hoatzin and seriema skeletons are also somewhat similar overall: I suppose you could believe that seriemas (and hence other cariamaens) are just big, long-legged hoatzins, modified for a cursorial, raptorial lifestyle whereas the various modifications possessed by hoatzins reflect their folivorous lifestyle (these features include a bizarre sternum where the keel is virtually absent anteriorly, thereby allowing room for the huge crop, and a notably deep, short pelvis) [the adjacent photos show, from top to bottom, a hoatzin skeleton, and a Cariama cristata skull and partial postcranium. Sorry the hoatzin photo is so bad]. Hoatzins and seriemas also possess a few bony and soft-tissue characters that are shared only by these birds, turacos and cuckoos: these include details of the hip musculature and the presence of distinctive bony recesses on the top of the pelvis.
Hoatzin and seriema skeletons are also somewhat similar overall: I suppose you could believe that seriemas (and hence other cariamaens) are just big, long-legged hoatzins, modified for a cursorial, raptorial lifestyle whereas the various modifications possessed by hoatzins reflect their folivorous lifestyle (these features include a bizarre sternum where the keel is virtually absent anteriorly, thereby allowing room for the huge crop, and a notably deep, short pelvis) [the adjacent photos show, from top to bottom, a hoatzin skeleton, and a Cariama cristata skull and partial postcranium. Sorry the hoatzin photo is so bad]. Hoatzins and seriemas also possess a few bony and soft-tissue characters that are shared only by these birds, turacos and cuckoos: these include details of the hip musculature and the presence of distinctive bony recesses on the top of the pelvis.

Olson (1985) supported the possible monophyly of the South American landbird group, writing ‘the seriemas and hoatzins appear to be part of an early radiation of primitive land birds, members of which have persisted in South America, perhaps as a result of its isolation’ (pp. 143-144). He further suggested that falcons ‘probably represent a raptorial branch of this radiation’ (p. 144), a suggestion presumably based on the anatomy of caracaras. Note that falconids are essentially a South American group (only a few, recently evolved genera have left the continent). A few fossil taxa might also be interpreted as providing support for the monophyly of this group. Mourer-Chauviré (1983) regarded idiornithids as similar to both seriemas and hoatzins and her conclusions are similar overall to those of Olson (1985). Incidentally, both Olson and Mourer-Chauviré came up with the same idea independently. A long delay in publication meant that Olson (1985) came out after Mourer-Chauviré (1983), by which time Olson decided not to rewrite his text: ‘partly out of laziness and frustration but more to show that we arrived independently at the same basic conclusions’ (p. 151).
 Among the most enigmatic of Cenozoic fossil birds is Foro panarium (image at left) from the Eocene Green River Formation of Wyoming: it seems to combine features of hoatzins, cuckoos and turacos but, interestingly, is also superficially raptor-like. This could be used to provide tentative support for Olson’s idea that falcons might be linked to hoatzins and other South American landbirds.
Among the most enigmatic of Cenozoic fossil birds is Foro panarium (image at left) from the Eocene Green River Formation of Wyoming: it seems to combine features of hoatzins, cuckoos and turacos but, interestingly, is also superficially raptor-like. This could be used to provide tentative support for Olson’s idea that falcons might be linked to hoatzins and other South American landbirds.
We saw above that hoatzins have often been linked with turacos, and that turacos share with hoatzins and seriemas that unusual and distinctive robust, bowed third metacarpal, as well as other characters. Might turacos also, then, be members of the South American landbird group? True, they aren’t South American but African (though with fossil representatives in
Based – it has to be said – on just a handful of detailed morphological characters, combined with some inference based on biogeography and superficial similarity, the South American landbird group theory suggests the following: that there might be a hoatzin-cariamaen clade, probably persisting as relicts in South America but more widespread during the early Cenozoic. It may perhaps involve turacos, and perhaps also falcons. Finally,
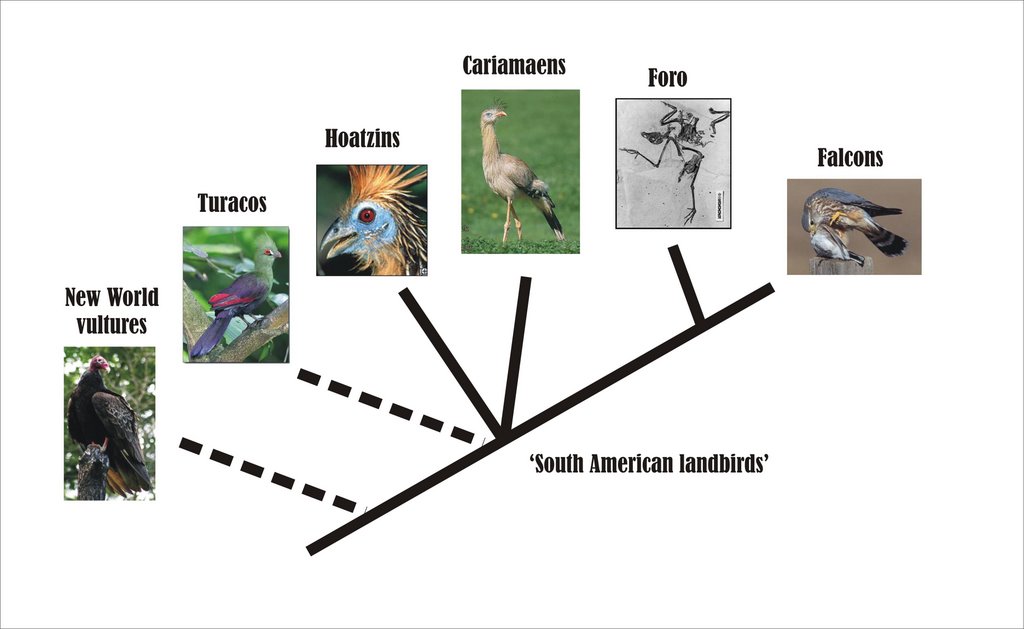
Predictably, I cannot help but find the idea that condors, caracaras, turacos and hoatzins are all close kin of phorusrhacids highly appealing, but it does all seem very vague and sadly lacking in good character support. So, do larger studies – those incorporating molecular and/or morphological information from lots of taxa – support a possible link between these birds? See the next post (Goodbye my giant predatory, cursorial, flightless hoatzin).
For the latest news on Tetrapod Zoology do go here.
Refs - -
Mayr, G. 2002. A new specimen of Salmila robusta (Aves: Gruiformes: Salmilidae n. fam.) from the Middle Eocene of Messel. Paläontologische Zeitschrift 76, 305-316.
Mourer-Chauviré, C. 1993. Les Gruiformes (Aves) des Phosphorites du Quercy (France). 1. Sous-ordre Cariamae (Cariamidae et Phorusrhacidae). Systématique et biostratigraphie. Palaeovertebrata 13, 83-143.
Olson, S. L. 1985. The fossil record of birds. In Avian Biology, Volume III, pp. 79-238.
Labels: birds, Cenozoic, phorusrhacids
posted by Darren Naish at 11:14 AM
15 comments
![]()
![]()